অধ্যায় ০৭ মৌলিক প্রক্রিয়াসমূহ
৭.১ জিনগত উপাদান হিসাবে ডিএনএ
আপনি পূর্ববর্তী অধ্যায়ে পড়েছেন যে বৈশিষ্ট্য বা চরিত্রগুলি জিনের মাধ্যমে পিতামাতা থেকে সন্তানদের মধ্যে বংশানুক্রমিক হয়। আপনি এও জানেন যে এই জিনগুলি ক্রোমোজোমে উপস্থিত থাকে যা নিউক্লিক অ্যাসিড এবং প্রোটিন দ্বারা গঠিত। তবে, বৈশিষ্ট্যের প্রকাশের জন্য দায়ী জিনের প্রকৃতি বোঝা বৈজ্ঞানিক সম্প্রদায়ের সামনে সবচেয়ে বড় চ্যালেঞ্জগুলির মধ্যে একটি ছিল। এই প্রশ্নের উত্তর এসেছিল কিছু পরীক্ষামূলক প্রমাণের পরে যা দেখায় যে ডিঅক্সিরাইবোনিউক্লিক অ্যাসিড (ডিএনএ) কিছু ভাইরাস ব্যতীত যেকোনো জীবের বৈশিষ্ট্য বা চরিত্র নির্ধারণ করে।
ডিএনএ আবিষ্কারের কৃতিত্ব জোহান ফ্রিডরিখ মিশারের, যিনি প্রথমবারের মতো পুঁজ কোষের নিউক্লিয়াস থেকে একটি অম্লীয় পদার্থ পৃথক করেছিলেন এবং নিউক্লিন নাম দিয়েছিলেন যাতে ডিএনএ এবং প্রোটিন ছিল। ক্রোমোজোম এবং নিউক্লিয়াসে উপস্থিতির কারণে এই দুটি রাসায়নিক উপাদান; নিউক্লিক অ্যাসিড (প্রধানত ডিএনএ) এবং প্রোটিন জিনগত উপাদান হওয়ার সম্ভাব্য প্রার্থী হয়ে ওঠে। তবুও, জিনগত উপাদানের প্রকৃতি দীর্ঘ সময়ের জন্য অজানা রয়ে গিয়েছিল। ধীরে ধীরে, বিভিন্ন গবেষক দ্বারা অণুজীবের সাথে পরীক্ষা-নিরীক্ষার ফলাফল প্রদান করে যা ডিএনএকে জিনগত উপাদান হিসাবে সমর্থন করে এমন প্রমাণ সরবরাহ করে।
৭.১.১ রূপান্তরকারী নীতির আবিষ্কার
১৯২৮ সালে, একজন ব্রিটিশ মেডিকেল অফিসার, ফ্রেডরিক গ্রিফিথ স্তন্যপায়ী প্রাণীতে ব্যাকটেরিয়া স্ট্রেপ্টোকক্কাস নিউমোনিয়া (যাকে ডিপ্লোকক্কাস নিউমোনিয়াও বলা হয়) দ্বারা সৃষ্ট নিউমোনিয়ার বিরুদ্ধে একটি ভ্যাকসিন তৈরির প্রক্রিয়ায় একটি পর্যবেক্ষণ করেছিলেন, যা মানুষের মধ্যে নিউমোনিয়া সৃষ্টি করে এবং সাধারণত ইঁদুরের মধ্যে প্রাণঘাতী। তিনি ব্যাকটেরিয়ার দুটি ভিন্ন স্ট্রেন (প্রকার) চিহ্নিত করেছিলেন, যেমন ভাইরুলেন্ট (রোগ সৃষ্টিকারী) যার কোষের চারপাশে একটি পলিস্যাকারাইড ক্যাপসুল রয়েছে এবং নন-ভাইরুলেন্ট (অনুপদ্রব)। ভাইরুলেন্ট স্ট্রেনে, প্রতিটি ব্যাকটেরিয়া একটি পলিস্যাকারাইড ক্যাপসুল দ্বারা বেষ্টিত থাকে যার কারণে ব্যাকটেরিয়ার কলোনি যখন একটি অ্যাগার প্লেটে জন্মায় তখন মসৃণ দেখায় এবং মসৃণ স্ট্রেন (এস) হিসাবে উল্লেখ করা হয়। নন-ভাইরুলেন্ট স্ট্রেনে পলিস্যাকারাইড আবরণের অভাব থাকে এবং রুক্ষ দেখতে কলোনি তৈরি করে এবং রুক্ষ স্ট্রেন (আর) হিসাবে উল্লেখ করা হয়। $\mathrm{S}$ প্রকারের ব্যাকটেরিয়া নিউমোনিয়া সৃষ্টি করে ইঁদুরকে মেরে ফেলে।
গ্রিফিথ $\mathrm{S}$ এবং $\mathrm{R}$ প্রকারের ব্যাকটেরিয়া নিয়ে一系列 পরীক্ষা করেছিলেন (চিত্র ৭.১)। যখন তিনি জীবিত $\mathrm{S}$ ব্যাকটেরিয়া ইঁদুরের মধ্যে ইনজেক্ট করেছিলেন, ইঁদুরগুলি নিউমোনিয়ায় আক্রান্ত হয়ে মারা যায়। তবে, যখন তিনি $\mathrm{R}$ প্রকারের ব্যাকটেরিয়া দিয়ে ইঁদুরকে সংক্রমিত করেছিলেন তখন ইঁদুরগুলি কোনো অসুস্থতা দেখায়নি। এই দুটি পরীক্ষার ফলাফল নিশ্চিত করে যে $\mathrm{S}$ প্রকারের ব্যাকটেরিয়ায় উপস্থিত পলিস্যাকারাইড আবরণটি আপাতদৃষ্টিতে ভাইরুলেন্সের জন্য প্রয়োজনীয় ছিল।
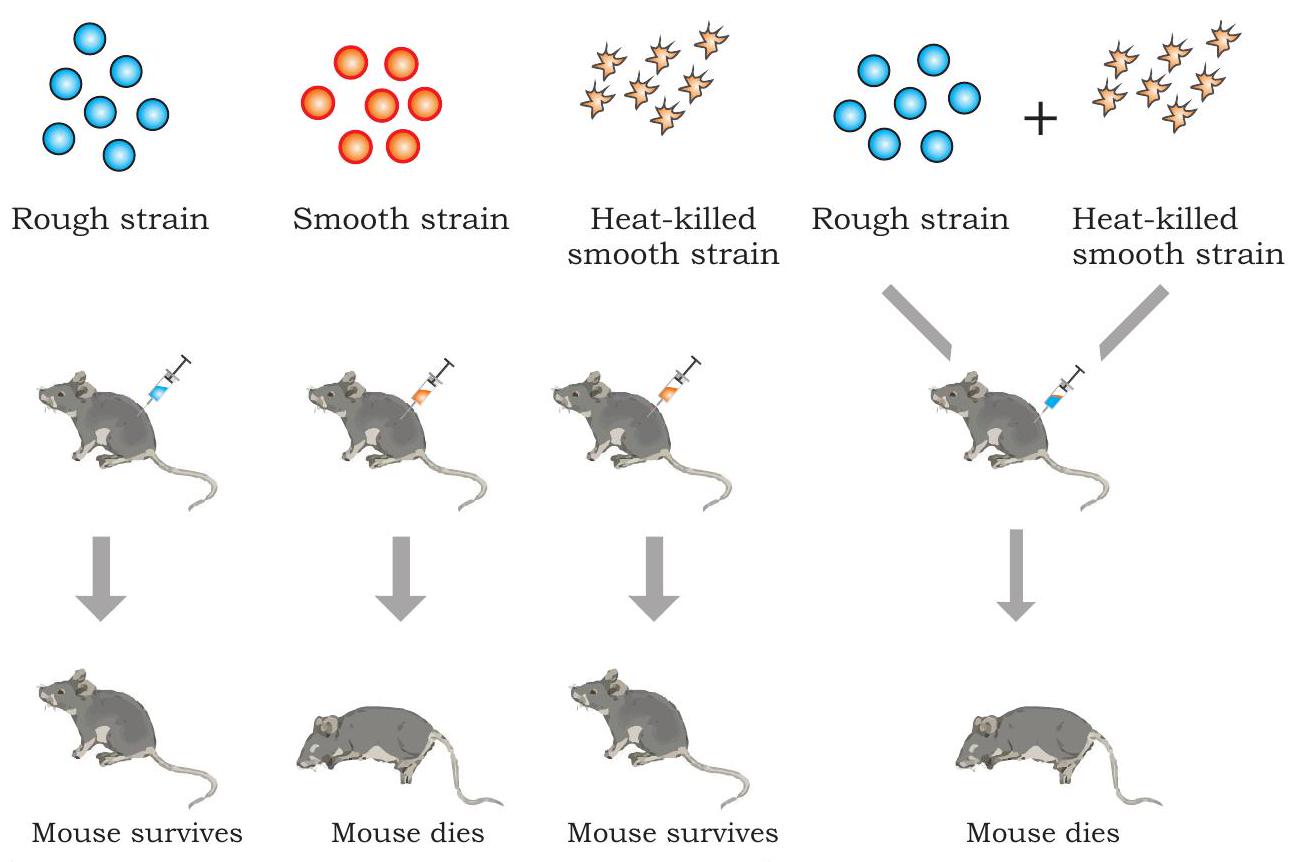
চিত্র ৭.১: গ্রিফিথের রূপান্তর পরীক্ষা
আরও বুঝতে, গ্রিফিথ কিছু ভাইরুলেন্ট $\mathrm{S}$ ব্যাকটেরিয়াকে সিদ্ধ করে মেরে ফেলেছিলেন এবং উক্ত তাপ-নিহত ব্যাকটেরিয়া ইঁদুরের মধ্যে ইনজেক্ট করেছিলেন। তার প্রত্যাশা অনুযায়ী, ইঁদুরগুলি বেঁচে গিয়েছিল। তবে, বেশ অপ্রত্যাশিতভাবে, যখন তাপ-নিহত $\mathrm{S}$ ব্যাকটেরিয়া এবং জীবিত $\mathrm{R}$ ব্যাকটেরিয়ার মিশ্রণ ইনজেক্ট করা হয়েছিল তখন ইঁদুরগুলি নিউমোনিয়ায় মারা যায়। মৃত ইঁদুরের রক্ত এবং টিস্যু তরল পরীক্ষা করে জীবিত $\mathrm{S}$ প্রকারের ব্যাকটেরিয়ার উপস্থিতি প্রকাশ পেয়েছে। উপরিউক্ত পর্যবেক্ষণের ভিত্তিতে, গ্রিফিথ সিদ্ধান্তে পৌঁছেছিলেন যে আর-স্ট্রেন ব্যাকটেরিয়াগুলি অবশ্যই তাপ-নিহত $S$ ব্যাকটেরিয়া থেকে যা তিনি ‘রূপান্তরকারী নীতি’ বলেছিলেন তা গ্রহণ করেছিল, যা তাদেরকে মসৃণ-আবরণযুক্ত ব্যাকটেরিয়ায় ‘রূপান্তরিত’ হতে এবং ভাইরুলেন্ট হতে দিয়েছে। তিনি এই ঘটনাটিকে রূপান্তর বলে অভিহিত করেছিলেন, যার অর্থ একটি কোষ থেকে অন্য কোষে জিনগত উপাদানের স্থানান্তর যা গ্রহীতা কোষের জিনগত গঠন পরিবর্তন করে। কিন্তু রূপান্তরকারী পদার্থের প্রকৃতি এখনও নির্ধারণ করা প্রয়োজন ছিল।
৭.১.২ রূপান্তরকারী নীতির জৈব রাসায়নিক চরিত্রায়ন
তিনজন বিজ্ঞানী, অসওয়াল্ড টি. অ্যাভেরি, কলিন ম্যাকলিওড এবং ম্যাকলিন ম্যাককার্টি গ্রিফিথের রূপান্তরকারী নীতি চিহ্নিত করার জন্য一系列 পরীক্ষা পরিচালনা করেছিলেন, এবং ১৯৪৪ সালে নিশ্চিত করা হয়েছিল যে রূপান্তরকারী এজেন্ট হল ডিএনএ (চিত্র ৭.২)। পরীক্ষার নকশায়, তারা ব্যাকটেরিয়ার মসৃণ স্ট্রেনের তিনটি প্রধান উপাদানের উপর ফোকাস করেছিল, যথা, ডিএনএ, আরএনএ এবং প্রোটিন। তারা ব্যাকটেরিয়ার মসৃণ স্ট্রেনের তাপ-নিহত একটি নির্যাস প্রস্তুত করেছিলেন যেখান থেকে লিপিড এবং কার্বোহাইড্রেট অপসারণ করা হয়েছিল। প্রোটিন, আরএনএ এবং ডিএনএ সম্বলিত নির্যাসের অবশিষ্ট উপাদানগুলি নির্যাসকে তিন ভাগে ভাগ করে আরও পরীক্ষার জন্য রাখা হয়েছিল। এই নির্যাসগুলিকে আলাদাভাবে হাইড্রোলাইটিক এনজাইম যেমন রাইবোনিউক্লিয়েজ (আরএনএজ), ডিঅক্সিরাইবোনিউক্লিয়েজ (ডিএনএজ) এবং প্রোটিজ দিয়ে চিকিত্সা করা হয়েছিল যথাক্রমে আরএনএ, ডিএনএ এবং প্রোটিনকে ক্ষয় করার জন্য, তাদের রূপান্তরকারী ক্ষমতা পরীক্ষা করার জন্য এনজাইম চিকিত্সিত প্রতিটি নির্যাসকে রুক্ষ স্ট্রেনের ব্যাকটেরিয়ার তিনটি ভিন্ন সংস্কৃতিতে স্থানান্তর করে। রুক্ষ স্ট্রেন থেকে মসৃণ স্ট্রেনে রূপান্তর সেই কলোনিগুলিতে পর্যবেক্ষণ করা হয়েছিল যেখানে আরএনএজ এবং প্রোটিজ চিকিত্সিত নির্যাস যোগ করা হয়েছিল এবং যেখানে ডিএনএজ চিকিত্সিত নির্যাস যোগ করা হয়েছিল সেই কলোনিতে নয়। এই ফলাফলগুলি কোনো সন্দেহ ছাড়াই প্রতিষ্ঠিত করে যে এটি ডিএনএ যা সম্ভাব্য রূপান্তরকারী নীতি হিসাবে কাজ করে।
হাইপোথিসিস: কোষগুলির জিনগত উপাদান হয় প্রোটিন বা নিউক্লিক অ্যাসিড (ডিএনএ বা আরএনএ)
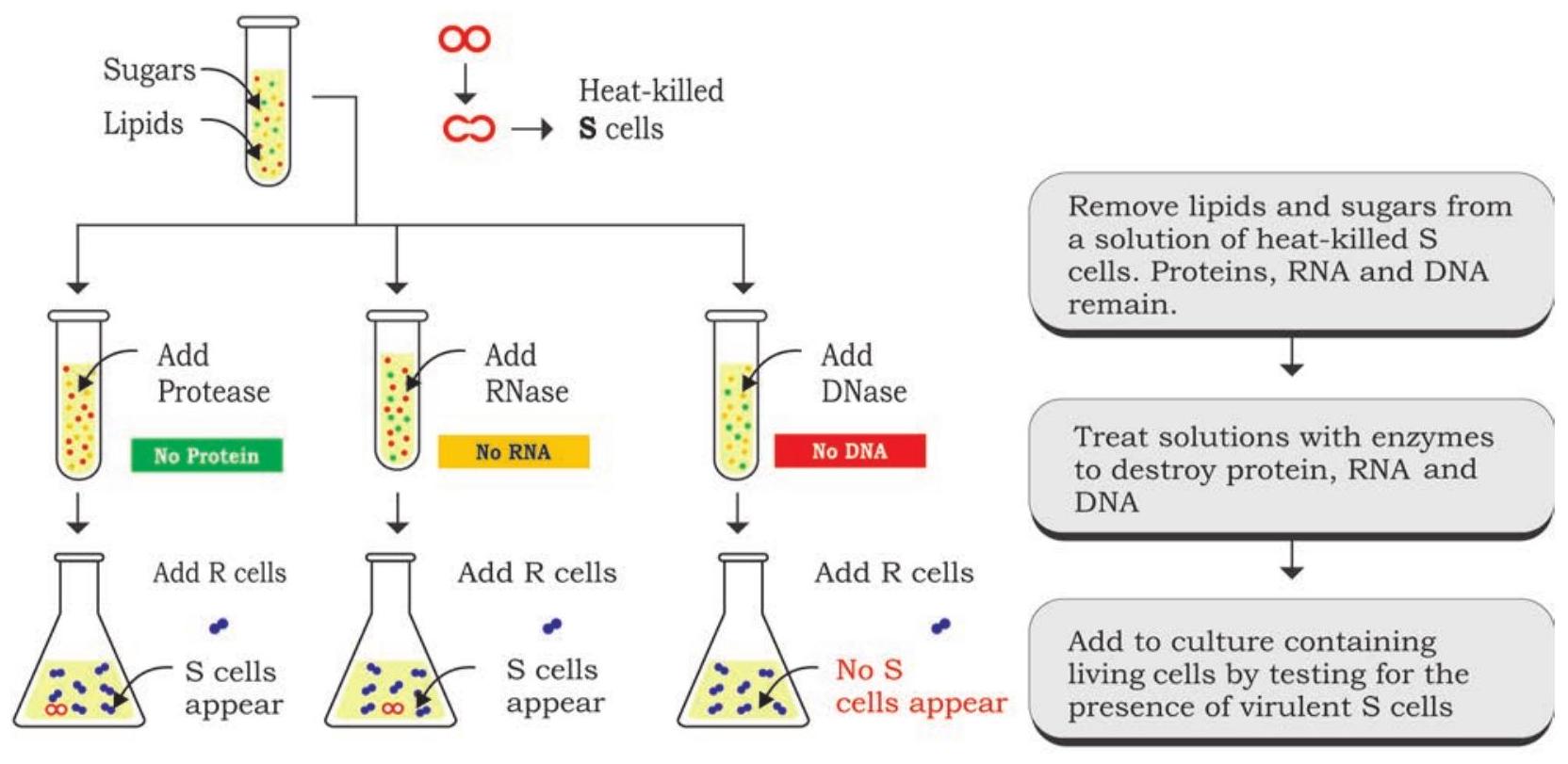
উপসংহার: রূপান্তরের জন্য ডিএনএ প্রয়োজন, তাই এটি কোষের জিনগত উপাদান
চিত্র ৭.২: রূপান্তরকারী নীতির নিশ্চিতকরণ
৭.১.৩ হার্শে - চেজ পরীক্ষা
পরে, আলফ্রেড হার্শে এবং মার্থা চেজ (১৯৫২) দ্বারা টি২ ব্যাকটেরিওফাজের সাথে পরিচালিত আরেকটি পরীক্ষা ডিএনএকে জিনগত উপাদান হিসাবে সমর্থন করে প্রমাণ প্রদান করে। ইশেরিচিয়া কোলাই ব্যাকটেরিয়াকে সংক্রমিত করে এমন ভাইরাস টি২ ব্যাকটেরিওফাজে প্রোটিন আবরণ দ্বারা বেষ্টিত ডিএনএ থাকে। যখন এটি একটি ব্যাকটেরিয়াল কোষকে সংক্রমিত করে, এটি বাইরের পৃষ্ঠে সংযুক্ত হয় এবং তারপর তার ডিএনএ কোষের মধ্যে ইনজেক্ট করে। টি২ ব্যাকটেরিওফাজ এবং ই. কোলাই-এর সাথে তাদের一系列 পরীক্ষায়, উদ্দেশ্য ছিল প্রতিষ্ঠা করা যে কোন উপাদান ফেজ কণার গুণনের জন্য দায়ী, ডিএনএ নাকি প্রোটিন। সহজে চিহ্নিত করার জন্য, $\mathrm{T} 2$ ব্যাকটেরিওফাজগুলি প্রাথমিকভাবে $E$. কোলাই-এর কলোনির সাথে পৃথকভাবে তেজস্ক্রিয় ফসফরাস $\left({ }^{32} \mathrm{P}\right)$ এবং তেজস্ক্রিয় সালফার $\left({ }^{35} \mathrm{S}\right)$ সম্বলিত মাঝারিতে জন্মানো হয়েছিল (চিত্র ৭.৩)। এর ফলে এক সেট ব্যাকটেরিওফাজ তেজস্ক্রিয় ফসফরাস $\left({ }^{32} \mathrm{P}\right)$ দিয়ে এবং অন্য সেট তেজস্ক্রিয় সালফার $\left({ }^{35} \mathrm{S}\right)$ দিয়ে লেবেল করা হয়েছিল।
${ }^{35} \mathrm{S}$ এবং ${ }^{32} \mathrm{P}$ লেবেলযুক্ত $\mathrm{T} 2$ ফেজগুলি এখন অলেবেলযুক্ত $E$. কোলাই ব্যাকটেরিয়াল কলোনির দুটি পৃথক সংস্কৃতিতে ইনোকুলেট করা হয়েছিল। সংক্রমণের পরে, ব্যাকটেরিয়াল কলোনিগুলিকে ব্যাকটেরিয়াল কোষগুলির বাইরের দিক থেকে কোনো অবশিষ্ট ফেজ এবং ফেজ অংশ অপসারণের জন্য একটি ব্লেন্ডারে নাড়ানো হয়েছিল। তারপর ব্লেন্ডারের মিশ্রণটি ব্যাকটেরিয়া (পেলেটে উপস্থিত) থেকে ফেজ ধ্বংসাবশেষ (সুপারনাট্যান্টে উপস্থিত) আলাদা করার জন্য সেন্ট্রিফিউজ করা হয়েছিল। যে ব্যাকটেরিয়াল সংস্কৃতির পেলেটগুলি তেজস্ক্রিয়তা দেখিয়েছিল সেগুলি তেজস্ক্রিয় ডিএনএ সম্বলিত ফেজ দ্বারা সংক্রমিত হয়েছিল, যেখানে, তেজস্ক্রিয়তা সুপারনাট্যান্টে পর্যবেক্ষণ করা হয়েছিল যা ${ }^{35} \mathrm{S}$ ব্যাকটেরিওফাজ দ্বারা সংক্রমিত হয়েছিল। এটি ইঙ্গিত করে যে প্রোটিনগুলি ফেজ থেকে ব্যাকটেরিয়ায় প্রবেশ করেনি। তাই, এই সিদ্ধান্তে উপনীত হওয়া হয়েছিল যে ব্যাকটেরিয়াল কোষে প্রবেশকারী উপাদান, অর্থাৎ, ডিএনএ জিনগত উপাদান হতে পারে।
যদিও উপরের পরীক্ষাগুলি ডিএনএকে জিনগত উপাদান হিসাবে সমর্থন করে শক্তিশালী প্রমাণ প্রদান করে, এটি স্পষ্ট ছিল না যে ডিএনএ অণুটি জিনগত তথ্যের ভাণ্ডার। এরউইন চারগাফ, মরিস উইলকিন্স, রোজালিন্ড ফ্রাঙ্কলিন, জেমস ওয়াটসন এবং ফ্রান্সিস ক্রিক দ্বারা পরবর্তী গবেষণাগুলি ডিএনএ কাঠামোর আবিষ্কারের দিকে নিয়ে যায়, স্পষ্ট করে যে কীভাবে ডিএনএ প্রচুর পরিমাণে তথ্য এনকোড করতে পারে (অধ্যায় ৩-এ বর্ণিত)।
$\hspace{3.5cm}$ব্যাকটেরিওফাজ

চিত্র ৭.৩: হার্শে-চেজ পরীক্ষা
৭.২ প্রোক্যারিওটিক এবং ইউক্যারিওটিক জিন সংগঠন
এটি ভালভাবে বোঝা যায় যে বৈশিষ্ট্যগুলি ‘জিন একক’ হিসাবে পিতামাতা থেকে সন্তানদের মধ্যে বংশানুক্রমিক হয় এবং কিছু ভাইরাস ব্যতীত সমস্ত জীবের মধ্যে ডিএনএ হল জিনগত উপাদান (যেখানে জিনগত উপাদান হল আরএনএ)। এটি জিনের সংগঠন সম্পর্কে প্রশ্ন উত্থাপন করে, এই সংগঠনটি প্রোক্যারিওট এবং ইউক্যারিওট উভয় ক্ষেত্রেই একই কিনা এবং এটি আণবিক স্তরে কীভাবে কাজ করে? জিন হল বংশগতির একক যা একটি নির্দিষ্ট বৈশিষ্ট্য বা চরিত্র নিয়ন্ত্রণ করে এবং অ্যালিল নামে পরিচিত বিকল্প রূপেও প্রকাশিত হতে পারে। অন্য কথায়, জিন হল ডিএনএ-এর একটি অংশ যা আরএনএ সংশ্লেষণের মাধ্যমে পলিপেপটাইড চেইনের সংশ্লেষণের মাধ্যমে নিজেকে প্রকাশ করে, যা আণবিক জীববিজ্ঞানের ‘কেন্দ্রীয় মতবাদ’ হিসাবে পরিচিত।
শুরুতে এটি প্রতিষ্ঠিত হয়েছিল যে বৈশিষ্ট্য বা চরিত্রগুলি ১৯০০ সালে মেন্ডেলের বংশগতির নীতির পুনঃআবিষ্কারের ভিত্তিতে জিন দ্বারা নিয়ন্ত্রিত হয়, এবং উদ্ভিদ এবং প্রাণী উভয় ক্ষেত্রে পরবর্তী一系列 গবেষণা এই সত্যটি প্রতিষ্ঠিত করে যে চরিত্র বা বৈশিষ্ট্যগুলি কিছু অন্তর্নিহিত নীতি দ্বারা নিয়ন্ত্রিত এবং নিয়ন্ত্রিত হয় যা এক প্রজন্ম থেকে অন্য প্রজন্মে স্থানান্তরিত হয়। বৈশিষ্ট্য বা চরিত্র নিয়ন্ত্রণকারী ফ্যাক্টর বা অন্তর্নিহিত একককে পরে ১৯০৯ সালে উইলহেম জোহানসেন ‘জিন’ নাম দিয়েছিলেন।
জিনের প্রকৃতি এবং কার্যকারিতা বোঝা বিংশ শতাব্দীর প্রথম দিকে বৈজ্ঞানিক সম্প্রদায়ের মূল ফোকাসগুলির মধ্যে একটি ছিল। ১৯৩০-এর দশকে জর্জ বিডল এবং এডওয়ার্ড ট্যাটামের ছাঁচ নিউরোস্পোরা ক্র্যাসার উপর কাজ একটি এনজাইমের সংশ্লেষণ নিয়ন্ত্রণে একটি জিনের সম্পর্ক প্রতিষ্ঠা করতে সাহায্য করেছিল।
ছাঁচ নিউরোস্পোরা ক্র্যাসার এই বৈশিষ্ট্য বিবেচনা করে যে এটি খুব সহজেই সাধারণ চিনি, অজৈব লবণ এবং ভিটামিন বায়োটিন সম্বলিত মাঝারিতে জন্মানো যায়, বিডল এবং ট্যাটাম এই ভিত্তিতে পরীক্ষা করেছিলেন যে জীবটি অন্যান্য প্রয়োজনীয় অ্যামিনো অ্যাসিড এবং নাইট্রোজেনাস বেস নিজেই সংশ্লেষণ করতে পারে (চিত্র ৭.৪)। এটি সুস্পষ্ট বলে বিবেচনা করা হয়েছিল যে সংশ্লেষণটি এনজাইম দ্বারা মধ্যস্থতা করা হয়, যা জিনগত নিয়ন্ত্রণে সংশ্লেষিত হয়। পরীক্ষার নকশা খুব সহজ ছিল যেখানে কনিডিয়া, অর্থাৎ, অযৌন বীজাণুগুলি মিউটেশন প্ররোচিত করার জন্য এক্স-রে দিয়ে বিকিরিত করা হয়েছিল (চিত্র ৭.৪)। বিকিরিত বীজাণু দ্বারা উৎপাদিত সন্তানদের কিছু নির্দিষ্ট ন্যূনতম মাঝারিতে জন্মানোর মাধ্যমে চিহ্নিত করা হয়েছিল। মিউটেশন সহ স্ট্রেন চিহ্নিত করার উদ্দেশ্যে, বিকিরিত বীজাণুর সন্তানদের বন্য প্রকারের সাথে ক্রস করা হয়েছিল এবং পরবর্তী সন্তানদের একটি নির্দিষ্ট অ্যামিনো অ্যাসিড বা ভিটামিনের জন্য ন্যূনতম মাঝারিতে জন্মানোর মাধ্যমে মিউট্যান্ট স্ট্রেন চিহ্নিত করা হয়েছিল (যে সংস্কৃতি মাঝারিতে একটি নির্দিষ্ট ভিটামিন বা অ্যামিনো অ্যাসিড ব্যতীত সমস্ত অ্যামিনো অ্যাসিড, নাইট্রোজেনাস বেস এবং ভিটামিন রয়েছে)। তাদের দ্বারা অনেক such মিউটেশন চিহ্নিত করা হয়েছিল এবং এটি জিনগতভাবে প্রতিষ্ঠিত হয়েছিল যে প্রতিটি মিউটেশন, আসলে, একটি নির্দিষ্ট এনজাইমের অ-কার্যকারিতার ফলাফল।
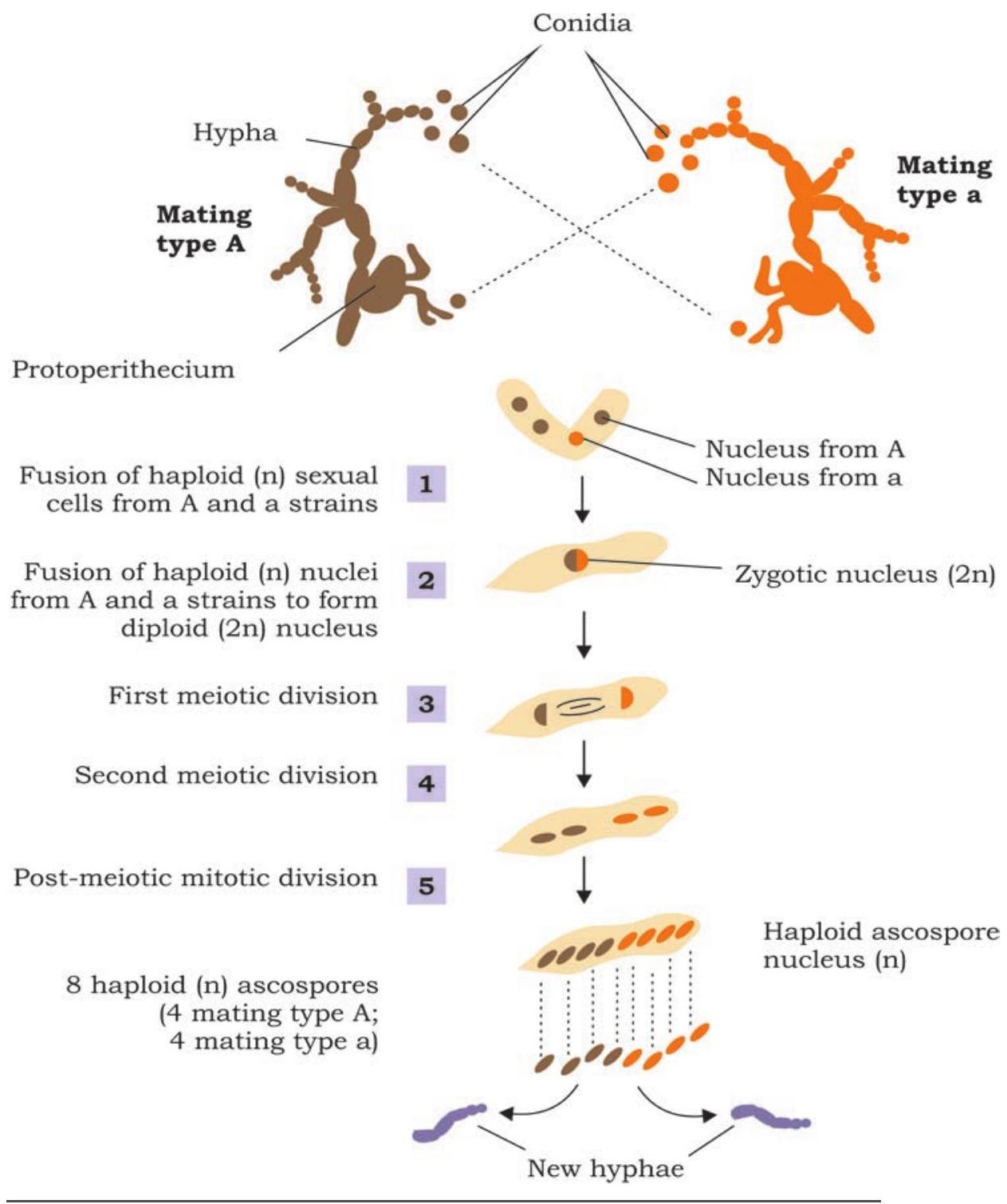
চিত্র ৭.৪: নিউরোস্পোরায় মিউটেশন সনাক্তকরণ দেখানো পরীক্ষা
পরবর্তীতে, এটি পর্যবেক্ষণ করা হয়েছিল যে সমস্ত প্রোটিন একক পলিপেপটাইড দ্বারা গঠিত নয় বরং একাধিক পলিপেপটাইড চেইন দ্বারা গঠিত। এই সত্য যে একটি জিন একটি পলিপেপটাইড এনকোড করে; কেন্দ্রীয় মতবাদটিও একটি জিন একটি প্রোটিন থেকে পরিবর্তিত হয়ে একটি জিন একটি পলিপেপটাইডে রূপান্তরিত হয়েছিল।
প্রায় একই সময়ে ১৯৪০-এর দশকের প্রথম দিকে, ক্রোমাটিন ফাইবারের সাইটোলজিকাল তদন্তে ইলেকট্রন মাইক্রোস্কোপির মাধ্যমে কিছুটা স্ট্রিংয়ের উপর পুঁতির মতো কাঠামো প্রকাশ পেয়েছিল (চিত্র ৭.৫) এবং এটি সহজেই সিদ্ধান্তে উপনীত হয়েছিল যে প্রতিটি পুঁতি সম্ভবত একটি জিনের প্রতিনিধিত্ব করে।

চিত্র ৭.৫: ক্রোমাটিনের স্ট্রিংয়ের উপর পুঁতির কাঠামো
পরবর্তী তদন্তে প্রকাশ পেয়েছে যে প্রতিটি পুঁতি একটি নিউক্লিওসোম (হিস্টোন অক্টামারের একটি কোর এবং $146 \mathrm{~bp}$ এর একটি ডাবল স্ট্র্যান্ডেড ডিএনএ ধারণ করে) এবং দুটি পুঁতির মধ্যে একটি স্ট্রিং, লিংকার ডিএনএ। এটিও প্রতিষ্ঠিত হয়েছিল যে প্রতিটি নিউক্লিওসোম তার লিংকার অঞ্চল সহ প্রায় $200 \mathrm{~bp}$ জড়িত। এটি একটি জিন হিসাবে বিবেচনা করা যায় না, কারণ অনেক ক্ষেত্রে জিনের আকার ২০০ নিউক্লিওটাইডের চেয়ে অনেক বড় হতে হবে। সহজ কারণ হল যে অনেক প্রোটিনের ১০০টিরও বেশি অ্যামিনো অ্যাসিড অবশিষ্টাংশ রয়েছে এবং তাদের সংশ্লিষ্ট নিয়ন্ত্রক জিনগুলি তার তিন গুণের কম হতে পারে না (কোডনের ট্রিপলেট প্রকৃতির ভিত্তিতে)।
এখন এটি স্পষ্ট হয়ে উঠেছে যে একটি জিন হল ডিএনএ-এর একটি অংশ যার নির্দিষ্ট প্রমোটার অঞ্চল রয়েছে, যেখানে আরএনএ পলিমারেজ বাঁধতে পারে এবং এমআরএনএ ট্রান্সক্রাইব করতে পারে। ট্রান্সক্রাইব করা এমআরএনএ তারপর অনুবাদের প্রক্রিয়ায় জড়িত হয়। প্রক্রিয়াটি ভাইরাস থেকে ব্যাকটেরিয়া, উদ্ভিদ এবং প্রাণী পর্যন্ত সমস্ত জীবের জন্য একই। একটি ভাইরাস, ব্যাকটেরিয়া বা উচ্চতর জীবের জিনগুলি তাদের গঠন এবং কার্যকারিতায় একই রকম? ক্রোমোজোমের একটি সম্পূর্ণ সেটে (সমস্ত জিন এবং ডিএনএ-এর অন্যান্য অংশ সহ) মোট ডিএনএ উপাদানকে জিনোম বলে। যেমন আমরা ভাইরাস বা ব্যাকটেরিয়ার ক্ষেত্রে দেখি, জিনোমের আকার তুলনামূলকভাবে ইউক্যারিওটিক জিনোমের চেয়ে অনেক ছোট। ইউক্যারিওটিক জিনোমগুলি প্রোক্যারিওটিক জিনোমের চেয়ে অনেক বেশি জটিল। উদ্ভিদ জিনোমগুলি অন্য যেকোনো ইউক্যারিওটিক জিনোমের চেয়ে আরও জটিল। জীবিত জীবের বিভিন্ন গোষ্ঠীতে জিনোমের আনুমানিক আকার চিত্র ৭.৬-এ দেখানো হয়েছে।
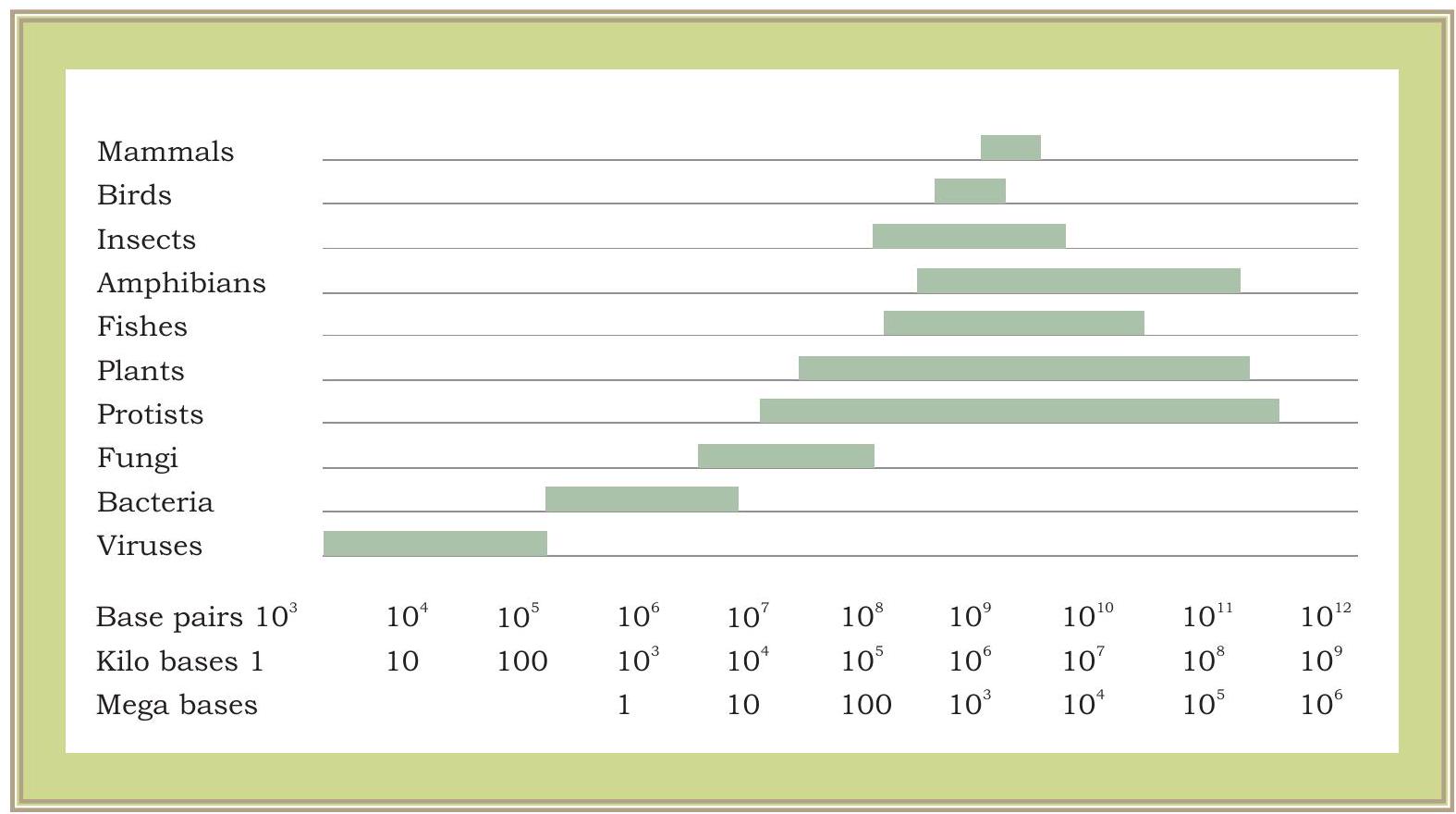
চিত্র ৭.৬: জীবিত জীবের বিভিন্ন গোষ্ঠীতে জিনোম আকারের তারতম্য। জিনোমের আকার হাজার হাজার নিউক্লিওটাইড জোড়ায় পরিমাপ করা হয়, অর্থাৎ, $1000 \mathrm{bp}=1$ কিলোবেস (কেবি) এবং ১০০০,০০০ $\mathrm{bp}=1000 \mathrm{~Kb}=1$ মেগাবেস (এমবি)
বেশিরভাগ ইউক্যারিওটে জিনোমের একটি বড় অংশ প্রকাশিত হয় না এবং নন-কোডিং সিকোয়েন্স হিসাবে থাকে। এটিও পর্যবেক্ষণ করা হয়েছে যে ইউক্যারিওটিক জিন এক্সপ্রেশনে পলিপেপটাইড চেইন সংশ্লেষণের প্রক্রিয়ায় জড়িত সক্রিয় এমআরএনএ-এর আকার প্রাথমিক ট্রান্সক্রিপ্টের চেয়ে অনেক ছোট। আসলে, অনেক ইউক্যারিওটিক জিন, যেমন, হিমোগ্লোবিনের $\beta$-গ্লোবিন জিন ট্রান্সক্রিপশনের পরে স্প্লাইসিং প্রক্রিয়ার মধ্য দিয়ে যায় যেখানে প্রাথমিক ট্রান্সক্রিপ্টের কয়েকটি বিচ্ছিন্ন অংশ অপসারণ করা হয় (ইনট্রন) এবং আরএনএ ট্রান্সক্রিপ্টের অবশিষ্ট অংশগুলি (এক্সন) এমআরএনএ গঠনের জন্য একত্রিত হয়।
একটি ইউক্যারিওটিক কোষে দুই ধরনের জিনোম রয়েছে: (i) নিউক্লিয়ার জিনোম এবং (ii) অর্গানেলার জিনোম।
নিউক্লিয়ার জিনোম
ডিএনএ-র বেশিরভাগ অংশ নিউক্লিয়াসে পাওয়া যায় এবং নিউক্লিয়ার ডিএনএ নামে পরিচিত। প্রোক্যারিওটে, বেশিরভাগ জিনোম কোডিং ডিএনএ সিকোয়েন্স দ্বারা গঠিত হয় যখন ইউক্যারিওটিক জিনোমে কোডিং অঞ্চলগুলি মোট জিনোমের তুলনামূলকভাবে খুব ছোট অংশ তৈরি করে। উদাহরণস্বরূপ, মানুষের জিনোমের আকার প্রায় $3,000 \mathrm{Mb}$ বা ৩ বিলিয়ন বেস পেয়ার ডিএনএ এবং ২০,০০০-এরও বেশি জিন থাকার অনুমান করা হয়, যা মোট জিনোমের প্রায় $2 %$ গঠন করে। জিনোমের ননকোডিং অঞ্চলে, এমন সিকোয়েন্স রয়েছে যা ট্যান্ডেম অ্যারের আকারে হাজার থেকে কয়েক মিলিয়ন বার পুনরাবৃত্তি হয়। জিনোমে এই পুনরাবৃত্তিমূলক ডিএনএ সিকোয়েন্সের আকার এবং সংখ্যা উল্লেখযোগ্যভাবে পরিবর্তিত হয়।
একটি ইউক্যারিওটিক কোষে নিউক্লিয়ার জিনোম ছোট ঘনীভূত এককে সংগঠিত হয়, যা রৈখিক ডিএনএ অণু ধারণকারী ক্রোমোজোম নামে পরিচিত। একটি ব্যক্তির হ্যাপ্লয়েড জিনোমে ক্রোমোজোমের একটি সম্পূর্ণ সেট অক্ষর n দ্বারা প্রতিনিধিত্ব করা হয়। বেশিরভাগ জীব প্রতিটি কোষে ক্রোমোজোমের দুটি সেট বহন করে এবং ডিপ্লয়েড জীব (2n) নামে পরিচিত।
অর্গানেলার জিনোম
নিউক্লিয়ার ডিএনএ ছাড়াও, কয়েকটি ঝিল্লি-বাউন্ড সেলুলার অর্গানেল যেমন ক্লোরোপ্লাস্ট এবং মাইটোকন্ড্রিয়া অর্গানেল-নির্দিষ্ট ডিএনএ ধারণ করে। অর্গানেল জিনোমগুলি বেশিরভাগই বৃত্তাকার ডাবল স্ট্র্যান্ডেড ডিএনএ এবং প্রতিটি অর্গানেলে একাধিক কপিতে উপস্থিত থাকে (চিত্র ৭.৭)। এগুলি সেমি-কনজারভেটিভ পদ্ধতিতে প্রতিলিপি করে এবং নিউক্লিয়ার জিনোম থেকে আলাদাভাবে বংশানুক্রমিক হয়। অর্গানেল ডিএনএতে এমন জিন থাকে যা অর্গানেল-নির্দিষ্ট কার্যকারিতার জন্য প্রয়োজনীয় এবং সাধারণত ইউনিপ্যারেন্টাল এবং মহিলা গ্যামেটের মাধ্যমে পরবর্তী প্রজন্মে বংশানুক্রমিক হয়।

চিত্র ৭.৭: অর্গানেলার ডিএনএ
৭.৩ ডিএনএ প্রতিলিপি
যখন ডিএনএ-এর ত্রিমাত্রিক কাঠামো ১৯৫৩ সালে জেমস ওয়াটসন এবং ফ্রান্সিস ক্রিক দ্বারা প্রস্তাবিত হয়েছিল, যে বৈশিষ্ট্যটি জীববিজ্ঞানীদের সবচেয়ে বেশি উত্তেজিত করেছিল তা হল দুটি পলিনিউক্লিওটাইড চেইনের বেসের পরিপূরক সম্পর্ক। ওয়াটসন এবং ক্রিক অবিলম্বে একটি মডেল প্রস্তাব করেছিলেন যে জিনগত তথ্য কপি করার ভিত্তি হল পরিপূরকতা (বিস্তারিত জানার জন্য ইউনিট II, অধ্যায় ২ দেখুন)। তাদের প্রস্তাবিত মডেল অনুসারে, ডিএনএ-এর দুটি স্ট্র্যান্ড প্রতিলিপির সময় পৃথক হয়; প্রতিটি স্ট্র্যান্ড বেস পেয়ারিং-এর নির্দিষ্টতার কারণে একটি নতুন পরিপূরক স্ট্র্যান্ডের সংশ্লেষণের জন্য একটি টেমপ্লেট হিসাবে কাজ করে (অর্থাৎ, অ্যাডেনিনের সাথে থাইমিন এবং গুয়ানিনের সাথে সাইটোসিন)। এইভাবে প্যারেন্টাল ডুপ্লেক্স ডিএনএ দুটি অভিন্ন ডটার ডুপ্লেক্স গঠন করে, যার প্রতিটিতে একটি প্যারেন্টাল স্ট্র্যান্ড এবং একটি নতুন সংশ্লেষিত ডটার স্ট্র্যান্ড থাকে। ডিএনএ প্রতিলিপির এই ফর্মটিকে সেমিকনজারভেটিভ প্রতিলিপি বলা হয় (চিত্র ৭.৮)। এই মোডের প্রতিলিপির প্রমাণ ১৯৫৮ সালে ম্যাথিউ মেসেলসন এবং ফ্রাঙ্কলিন স্টাহল প্রদান করেছিলেন।
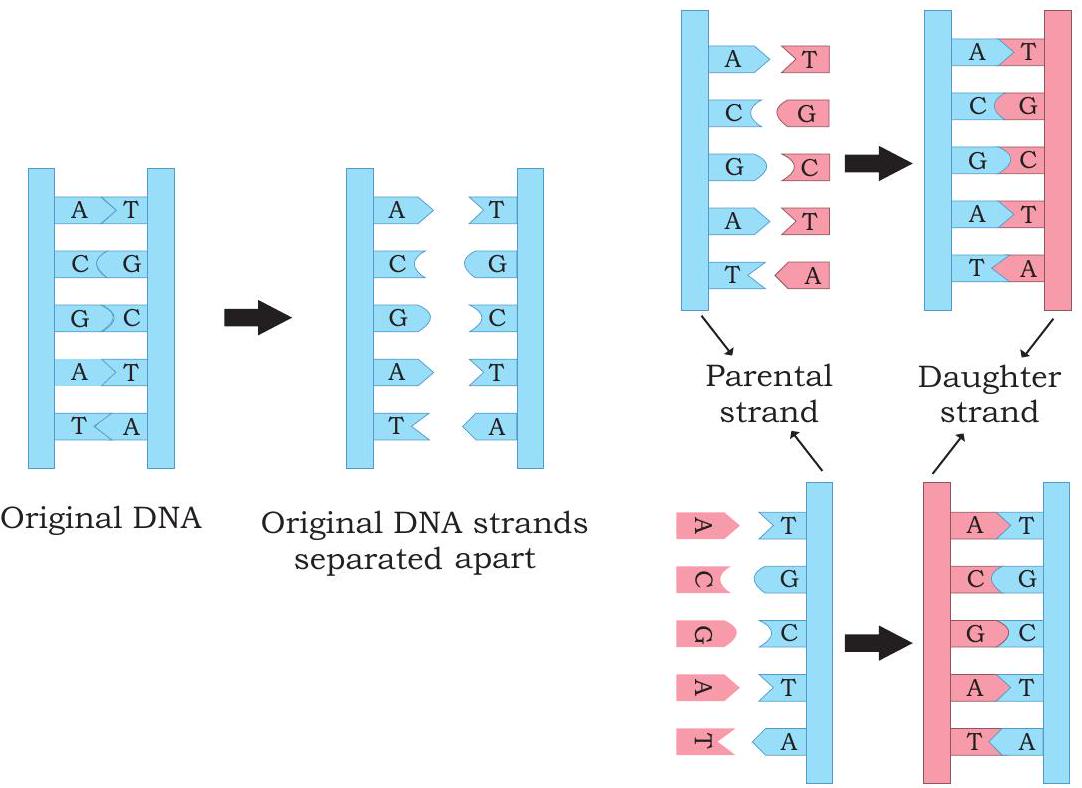
চিত্র ৭.৮: ডিএনএ প্রতিলিপির সেমিকনজারভেটিভ মোড
৭.৩.১ মেসেলসন এবং স্টাহল পরীক্ষা
পুরানো এবং নতুন স্ট্র্যান্ডের মধ্যে পার্থক্য করতে, মেসেলসন এবং স্টাহল নাইট্রোজেনের দুটি আইসোটোপ ব্যবহার করেছিলেন, ${ }^{14} \mathrm{~N}$ (সাধারণ ফর্ম) এবং ${ }^{15} \mathrm{~N}$ (একটি বিরল, ভারী ফর্ম)। তারা $E$. কোলাই ব্যাকটেরিয়াকে একমাত্র নাইট্রোজেন উৎস হিসাবে ভারী আইসোটোপ সম্বলিত একটি মাঝারিতে জন্মিয়েছিল। কয়েক প্রজন্মের পরে, সমস্ত ই. কোলাই কোষে ডিএনএ-এর পিউরিন এবং পাইরিমিডিন বেসে ${ }^{15} \mathrm{~N}$ অন্তর্ভুক্ত ছিল। ঘনত্ব গ্রেডিয়েন্ট সেন্ট্রিফিউগেশন ব্যবহার করে, তারা পর্যবেক্ষণ করেছিল যে $E$. কোলাই থেকে নিষ্কাশিত ডিএনএ ${ }^{15} \mathrm{~N}$-এ জন্মানোর সময় সেন্ট্রিফিউজ টিউবের নীচের দিকে একটি একক ব্যান্ড তৈরি করেছিল, যখন ${ }^{14} \mathrm{~N}$ মাঝারিতে জন্মানো ব্যাকটেরিয়াল কোষ থেকে নিষ্কাশিত ডিএনএ শীর্ষের কাছাকাছি একটি ব্যান্ড গঠন করেছিল। এটি ইঙ্গিত করে যে $E$. কোলাই কোষের ডিএনএ ${ }^{15} \mathrm{~N}$ মাঝারিতে জন্মানোর সময় হালকা আইসোটোপ $\left({ }^{14} \mathrm{~N}\right)$ সম্বলিত মাঝারিতে জন্মানো ব্যাকটেরিয়ার চেয়ে ঘন ছিল (চিত্র ৭.৯)।
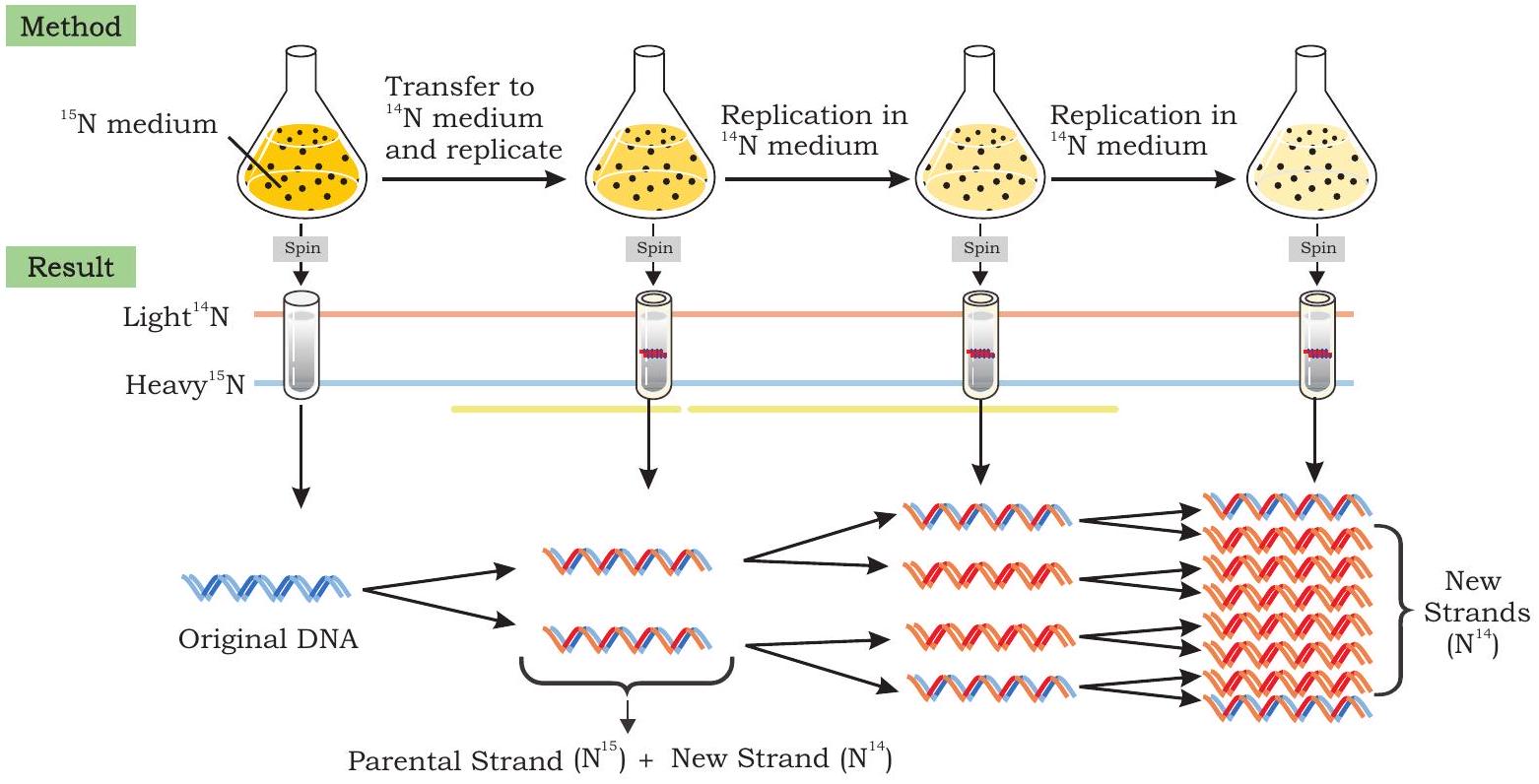
চিত্র ৭.৯: ডিএনএ প্রতিলিপির সেমিকনজারভেটিভ মোড নিশ্চিত করার জন্য মেসেলসন - স্টাহল পরীক্ষা
মেসেলসন এবং স্টাহল তারপর ই. কোলাই ব্যাকটেরিয়াকে ${ }^{15} \mathrm{~N}$ মাঝারি থেকে ${ }^{14} \mathrm{~N}$ মাঝারিতে স্থানান্তরিত করেছিলেন এবং ব্যাকটেরিয়াল কোষগুলি গুণিত হওয়ার সাথে সাথে বিভিন্ন সময়ের ব্যবধানে ডিএনএ সংগ্রহ করেছিলেন $(E$. কোলাই প্রতি ২০ মিনিট পরে বিভাজিত হয়)। প্রথম রাউন্ড বিভাজনের পরে $E$. কোলাই কোষ থেকে নিষ্কাশিত ডিএনএ (প্রজন্ম I) যখন সিজিয়াম ক্লোরাইড লবণ ব্যবহার করে ঘনত্ব গ্রেডিয়েন্ট সেন্ট্রিফিউগেশন দ্বারা বিশ্লেষণ করা হয়েছিল) একটি একক ব্যান্ড তৈরি করেছিল, কিন্তু ভারী ডিএনএ $\left({ }^{15} \mathrm{~N}\right)$ এবং হালকা ডিএনএ $\left({ }^{14} \mathrm{~N}\right)$ ব্যান্ডের মধ্যবর্তী অবস্থানে।

ম্যাথিউ মেসেলসন এবং ফ্রাঙ্কলিন স্টাহল ডিএনএ প্রতিলিপির সেমি-কনজারভেটিভ মোড নিশ্চিত করেছিলেন
যখন ডিএনএ $E$. কোলাই কোষ থেকে নিষ্কাশিত করা হয়েছিল, ${ }^{14} \mathrm{~N}$ মাঝারিতে দ্বিতীয় রাউন্ড প্রতিলিপির পরে (প্রজন্ম II) সেন্ট্রিফিউজ টিউবে সমান তীব্রতার দুটি ব্যান্ড উপস্থিত হয়েছিল, একটি মধ্যবর্তী অবস্থানে এবং অন্যটি শুধুমাত্র ${ }^{14} \mathrm{~N}$ ডিএনএ ধারণকারী ডিএনএ-এর প্রত্যাশিত অবস্থানে। যখন অতিরিক্ত রাউন্ড প্রতিলিপির পরে ডিএনএ-এর নমুনা সংগ্রহ করা হয়েছিল তখন তারা দুটি ব্যান্ড তৈরি করেছিল। হালকা ডিএনএ-এর প্রতিনিধিত্বকারী ব্যান্ডগুলি ক্রমাগত পুরু হয়ে উঠছিল কিন্তু মধ্যবর্তী অবস্থানে ব্যান্ডটি অপরিবর্তিত রয়ে গিয়েছিল।
বক্স ১
ঘনত্ব গ্রেডিয়েন্ট সেন্ট্রিফিউগেশন
এটি তাদের ঘনত্বের ভিত্তিতে একটি মিশ্রণ থেকে অণুগুলিকে পৃথক করার জন্য একটি সেন্ট্রিফিউগেশন কৌশল। সেন্ট্রিফিউগেশন টিউবটি সিজিয়াম ক্লোরাইড ($\mathrm{CsCl}$) এর একটি ভারী লবণ দ্রবণ এবং যার ঘনত্ব পরিমাপ করতে হবে তার নমুনা দিয়ে পূর্ণ করা হয়। তারপর সেন্ট্রিফিউজ টিউবটিকে একটি আল্ট্রাসেন্ট্রিফিউজে কয়েক দিনের জন্য খুব উচ্চ গতিতে ঘোরানো হয়। আল্ট্রাসেন্ট্রিফিউজ দ্বারা উৎপন্ন বিশাল কৃত্রিম বল সিএস আয়নগুলিকে টিউবের নীচের দিকে স্থানান্তরিত করে, নীচে উচ্চ ঘনত্ব এবং শীর্ষে কম ঘনত্ব সহ একটি গ্রেডিয়েন্ট তৈরি করে। ডিএনএ স্ট্র্যান্ডগুলি লবণের ঘনত্বের সাথে মিল না হওয়া পর্যন্ত গ্রেডিয়েন্টে ভাসে বা ডুবে যায়।

৭.৩.২ মেসেলসন এবং স্টাহল দ্বারা ফলাফলের ব্যাখ্যা
প্রতিলিপির প্রথম রাউন্ডের পরে, প্রতিটি ডটার ডিএনএ ডুপ্লেক্স একটি হাইব্রিড ছিল যাতে প্যারেন্ট থেকে ${ }^{15} \mathrm{~N}$ ধারণকারী একটি ভারী স্ট্র্যান্ড এবং মাঝারি থেকে ${ }^{14} \mathrm{~N}$ ধারণকারী একটি হালকা স্ট্র্যান্ড ছিল। যখন এই হাইব্রিড ডুপ্লেক্স প্রতিলিপি হয়েছিল, ভারী টেমপ্লেট স্ট্র্যান্ডটি আরেকটি হাইব্রিড ডুপ্লেক্স ডিএনএ গঠন করেছিল যখন হালকা স্ট্র্যান্ডটি হালকা ডিএনএ ডুপ্লেক্স গঠন করেছিল। এইভাবে, মেসেলসন এবং স্টাহল পরীক্ষা স্পষ্টভাবে ওয়াট