அத்தியாயம் 07 அடிப்படை செயல்முறைகள்
7.1 மரபணுப் பொருளாக DNA
முந்தைய அத்தியாயத்தில் நீங்கள் படித்தபடி, பண்புகள் அல்லது குணங்கள் மரபணுக்கள் மூலம் பெற்றோரிடமிருந்து சந்ததிகளுக்கு பரம்பரையாக கடத்தப்படுகின்றன. இந்த மரபணுக்கள் குரோமோசோம்களில் அமைந்துள்ளன என்பதும் உங்களுக்குத் தெரியும், அவை நியூக்ளிக் அமிலங்கள் மற்றும் புரதங்களால் ஆனவை. இருப்பினும், ஒரு பண்பின் வெளிப்பாட்டிற்குக் காரணமான மரபணுவின் இயல்பைப் புரிந்துகொள்வது அறிவியல் சமூகத்தின் முன்னால் இருந்த மிகப்பெரிய சவால்களில் ஒன்றாகும். டி ஆக்ஸிரைபோ நியூக்ளிக் அமிலம் (DNA) சில வைரஸ்களைத் தவிர மற்ற அனைத்து உயிரினங்களின் பண்பு அல்லது அம்சத்தையும் தீர்மானிக்கிறது என்பதற்கான சில சோதனை ஆதாரங்களுக்குப் பிறகு இந்தக் கேள்விக்கான பதில் கிடைத்தது.
DNAயின் கண்டுபிடிப்புக்கு ஜோஹான் பிரெட்ரிக் மீஷ்சர் கிரெடிட் பெற்றார், அவர் முதன்முதலில் சீழ் செல்களின் கருக்களிலிருந்து ஒரு அமிலப் பொருளைத் தனிமைப்படுத்தி, DNA மற்றும் புரதத்தைக் கொண்ட நியூக்ளின் என்று பெயரிட்டார். குரோமோசோம் மற்றும் கருவில் இருப்பதால் இந்த இரண்டு வேதியியல் கூறுகள்; நியூக்ளிக் அமிலம் (முக்கியமாக DNA) மற்றும் புரதம் மரபணுப் பொருளாக இருக்கக்கூடிய சாத்தியமான வேட்பாளர்களாக மாறியது. இன்னும், மரபணுப் பொருளின் இயல்பு நீண்ட காலமாக அறியப்படாமலேயே இருந்தது. படிப்படியாக, வெவ்வேறு ஆராய்ச்சியாளர்களால் நுண்ணுயிரிகளுடன் செய்யப்பட்ட சோதனைகள் DNA மரபணுப் பொருளாக இருப்பதற்கு ஆதாரங்களை வழங்கும் முடிவுகளைத் தந்தன.
7.1.1 உருமாற்றக் கோட்பாட்டின் கண்டுபிடிப்பு
1928 ஆம் ஆண்டில், ஒரு பிரிட்டிஷ் மருத்துவ அதிகாரியான பிரெடெரிக் கிரிஃபித், பாலூட்டிகளில் Streptococcus pneumoniae (இது Diplococcus pneumoniae என்றும் அழைக்கப்படுகிறது) என்ற பாக்டீரியாவால் ஏற்படும் நிமோனியாவுக்கு எதிரான தடுப்பூசியை உருவாக்கும் போது ஒரு கவனிப்பைச் செய்தார், இது மனிதர்களில் நிமோனியாவை ஏற்படுத்துகிறது மற்றும் பொதுவாக சுண்டெலிகளில் கொடியது. அவர் பாக்டீரியத்தின் இரண்டு வெவ்வேறு இனங்களை (வகைகள்) அடையாளம் கண்டார், அதாவது நோயை உண்டாக்கும் (நோயை உண்டாக்கும்) செல்லைச் சுற்றி பாலிசாக்கரைடு உறை கொண்டது மற்றும் நோயற்றது (தீங்கற்றது). நோயை உண்டாக்கும் இனத்தில், ஒவ்வொரு பாக்டீரியமும் ஒரு பாலிசாக்கரைடு உறையால் சூழப்பட்டிருக்கும், இதன் காரணமாக அகர் தட்டில் வளரும் போது பாக்டீரியா காலனி மென்மையாகத் தோன்றும் மற்றும் மென்மையான இனம் (S) என்று குறிப்பிடப்படுகிறது. நோயற்ற இனத்தில் பாலிசாக்கரைடு பூச்சு இல்லை மற்றும் கரடுமுரடான தோற்றத்தைக் கொண்ட காலனியை உருவாக்குகிறது மற்றும் கரடுமுரடான இனம் (R) என்று குறிப்பிடப்படுகிறது. $\mathrm{S}$ வகை பாக்டீரியாக்கள் நிமோனியாவை ஏற்படுத்தி சுண்டெலிகளைக் கொல்கின்றன.
கிரிஃபித் $\mathrm{S}$ மற்றும் $\mathrm{R}$ வகை பாக்டீரியாக்களுடன் (படம் 7.1) தொடர் சோதனைகளைச் செய்தார். அவர் உயிர்ப்பான $\mathrm{S}$ பாக்டீரியாக்களை சுண்டெலிகளுக்குள் செலுத்தியபோது, சுண்டெலிகள் நிமோனியாவால் பாதிக்கப்பட்டு இறந்தன. இருப்பினும், அவர் $\mathrm{R}$ வகை பாக்டீரியாக்களால் சுண்டெலிகளைப் பாதித்தபோது, சுண்டெலிகள் எந்தக் கேடும் காட்டவில்லை. இந்த இரண்டு சோதனைகளின் முடிவுகள் $\mathrm{S}$ வகை பாக்டீரியாக்களில் இருந்த பாலிசாக்கரைடு உறை வெளிப்படையாக நோய்த்தன்மைக்கு அவசியம் என்பதை உறுதிப்படுத்தியது.
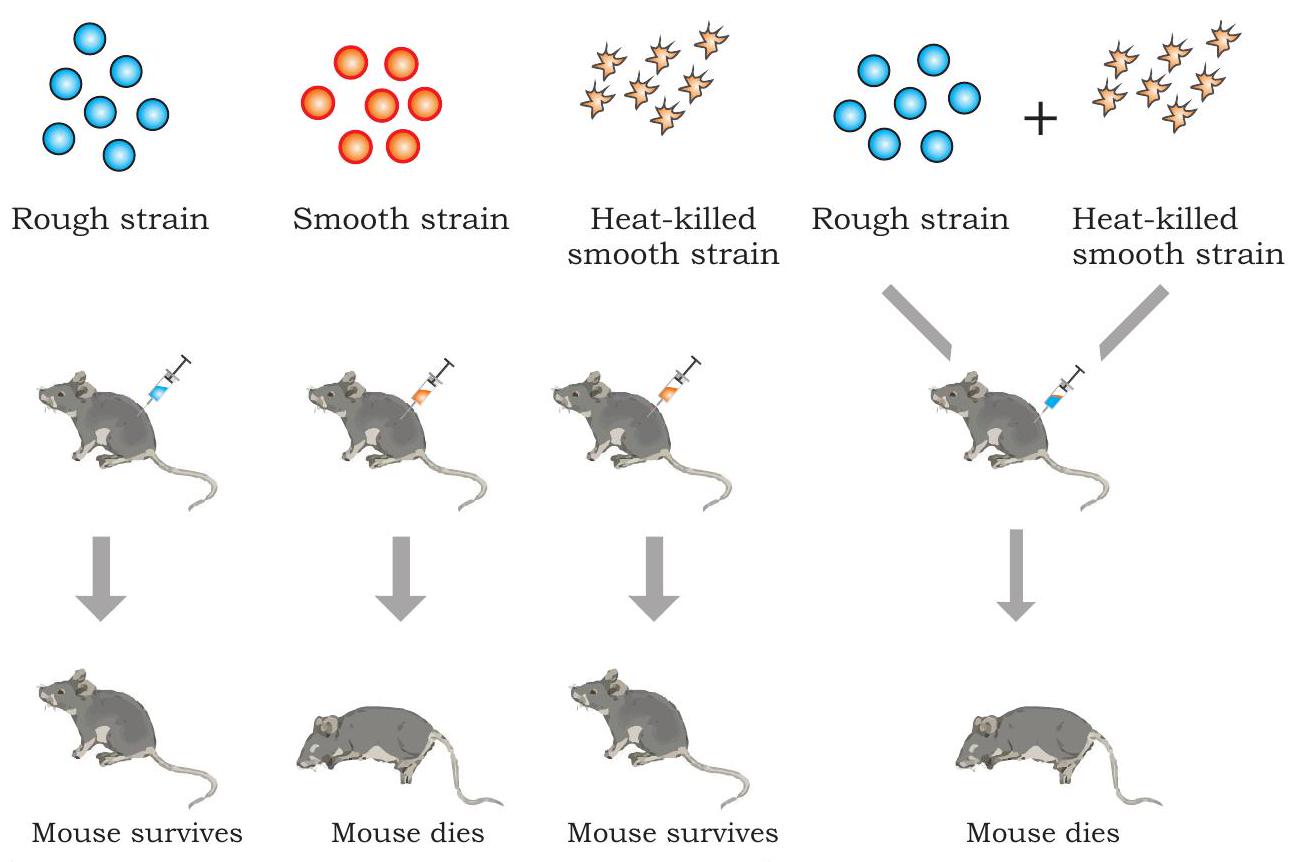
படம் 7.1: கிரிஃபித்தின் உருமாற்றச் சோதனை
மேலும் புரிந்துகொள்வதற்காக, கிரிஃபித் சில நோயை உண்டாக்கும் $\mathrm{S}$ பாக்டீரியாக்களை கொதிக்க வைத்துக் கொன்று, கூறப்பட்ட வெப்பத்தால் கொல்லப்பட்ட பாக்டீரியாக்களை சுண்டெலிகளுக்குள் செலுத்தினார். அவரது எதிர்பார்ப்புகளின்படி, சுண்டெலிகள் உயிர் பிழைத்தன. இருப்பினும், மிகவும் எதிர்பாராத விதமாக, வெப்பத்தால் கொல்லப்பட்ட $\mathrm{S}$ பாக்டீரியாக்கள் மற்றும் உயிர்ப்பான $\mathrm{R}$ பாக்டீரியாக்களின் கலவையை செலுத்தியபோது சுண்டெலிகள் நிமோனியாவால் இறந்தன. இறந்த சுண்டெலிகளின் இரத்தம் மற்றும் திசு திரவத்தின் பரிசோதனை உயிர்ப்பான $\mathrm{S}$ வகை பாக்டீரியாக்களின் இருப்பை வெளிப்படுத்தியது. மேலே உள்ள கவனிப்பின் அடிப்படையில், கிரிஃபித் R-இன பாக்டீரியாக்கள் வெப்பத்தால் கொல்லப்பட்ட $S$ பாக்டீரியாக்களிலிருந்து ‘உருமாற்றக் கோட்பாடு’ என அழைக்கப்படுவதை எடுத்துக்கொண்டிருக்க வேண்டும் என்று முடிவு செய்தார், இது அவற்றை மென்மையான பூச்சு பாக்டீரியாக்களாக ‘மாற்ற’ அனுமதித்து நோயை உண்டாக்கும் தன்மையைப் பெறச் செய்தது. அவர் இந்த நிகழ்வை உருமாற்றம் என்று அழைத்தார், இது ஒரு செல்லிலிருந்து மற்றொரு செல்லுக்கு மரபணுப் பொருளை மாற்றுவதைக் குறிக்கிறது, இது பெறும் செல்லின் மரபணு அமைப்பை மாற்றுகிறது. ஆனால் உருமாற்றப் பொருளின் இயல்பு இன்னும் தீர்மானிக்கப்பட வேண்டியிருந்தது.
7.1.2 உருமாற்றக் கோட்பாட்டின் உயிர்வேதியியல் பண்புக் கூறு
மூன்று விஞ்ஞானிகள், ஆஸ்வால்ட் டி. அவெரி, கோலின் மெக்லியோட் மற்றும் மெக்லின் மெக்கார்ட்டி ஆகியோர் கிரிஃபித்தின் உருமாற்றக் கோட்பாட்டை அடையாளம் காண தொடர் சோதனைகளை நடத்தினர், மேலும் 1944 ஆம் ஆண்டில் உருமாற்றக் காரணி DNA என்பது உறுதிப்படுத்தப்பட்டது (படம் 7.2). சோதனையின் வடிவமைப்பில், அவர்கள் பாக்டீரியாவின் மென்மையான இனத்தின் மூன்று முக்கிய கூறுகளில் கவனம் செலுத்தினர், அதாவது DNA, RNA மற்றும் புரதம். அவர்கள் வெப்பத்தால் கொல்லப்பட்ட பாக்டீரியாவின் மென்மையான இனத்தின் சாற்றைத் தயாரித்தனர், அதிலிருந்து கொழுப்புகள் மற்றும் கார்போஹைட்ரேட்டுகள் அகற்றப்பட்டன. புரதங்கள், RNA மற்றும் DNA ஆகியவற்றைக் கொண்ட சாற்றின் மீதமுள்ள கூறுகள் மேலும் சோதனைக்காக சாற்றை மூன்று பகுதிகளாகப் பிரிப்பதன் மூலம் தக்க வைக்கப்பட்டன. இந்தச் சாறுகள் தனித்தனியாக ரைபோநியூக்ளியேஸ் (RNase), டி ஆக்ஸிரைபோநியூக்ளியேஸ் (DNase) மற்றும் புரோட்டீஸ் போன்ற நீராற்பகுப்பு நொதிகளால் சிகிச்சையளிக்கப்பட்டன, முறையே RNA, DNA மற்றும் புரதத்தை சிதைக்க, அவற்றின் உருமாற்ற திறனுக்காக நொதி சிகிச்சையளிக்கப்பட்ட ஒவ்வொரு சாற்றையும் பாக்டீரியாவின் கரடுமுரடான இனத்தின் மூன்று வெவ்வேறு கலாச்சாரங்களுக்கு மாற்றுவதன் மூலம். RNase மற்றும் புரோட்டீஸ் சிகிச்சையளிக்கப்பட்ட சாறு சேர்க்கப்பட்ட காலனிகளில் கரடுமுரடான இனம் மென்மையான இனமாக மாற்றப்பட்டது மற்றும் DNase சிகிச்சையளிக்கப்பட்ட சாறு சேர்க்கப்பட்ட காலனியில் இல்லை. இந்த முடிவுகள் DNA தான் உருமாற்றக் கோட்பாடாக செயல்படுகிறது என்பதை சந்தேகத்திற்கு இடமின்றி நிறுவியது.
கருதுகோள்: செல்களின் மரபணுப் பொருள் புரதம் அல்லது நியூக்ளிக் அமிலம் (DNA அல்லது RNA) ஆகும்.
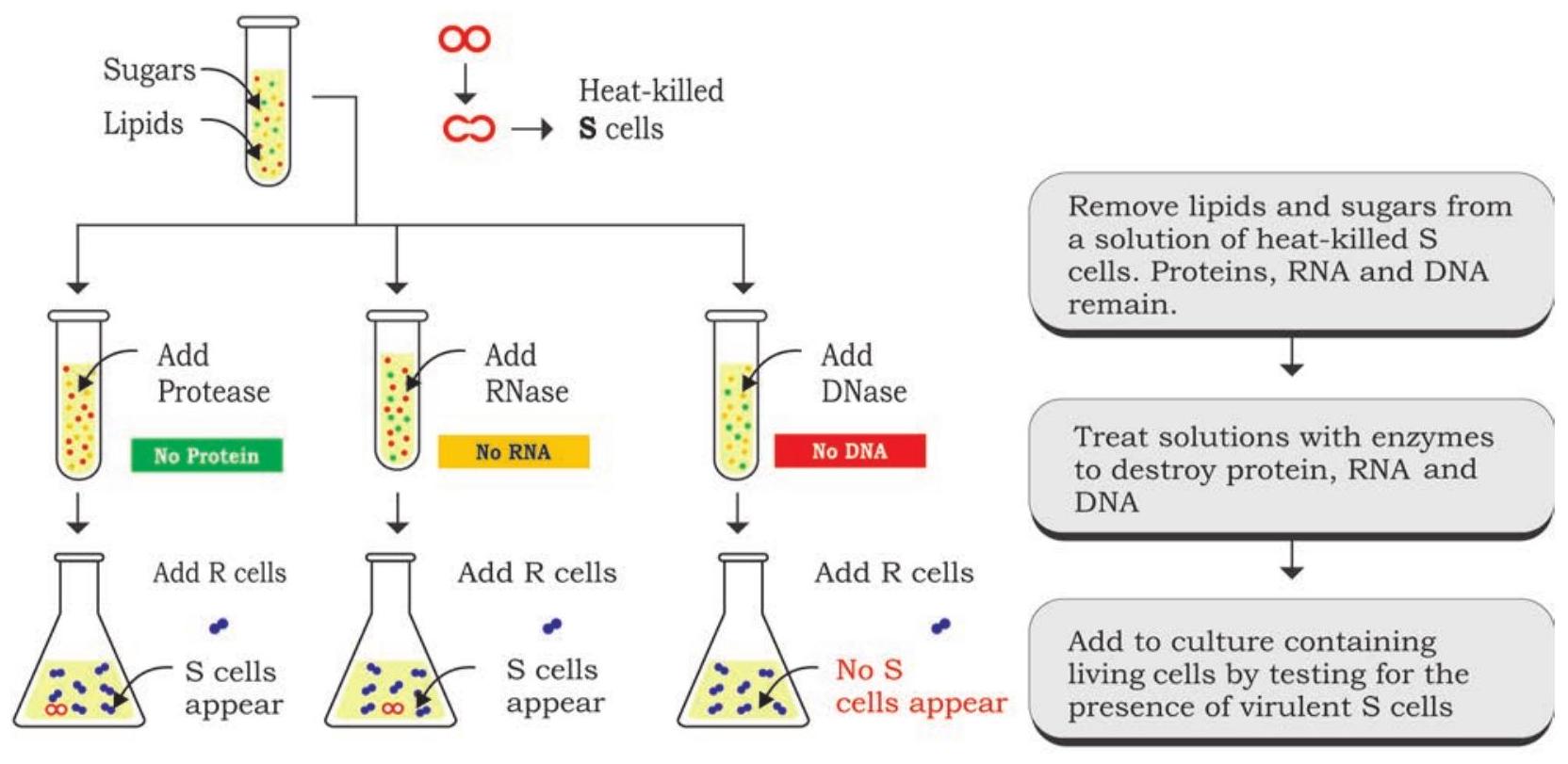
முடிவு: உருமாற்றத்திற்கு DNA தேவைப்படுகிறது, எனவே இது செல்லின் மரபணுப் பொருளாகும்.
படம் 7.2: உருமாற்றக் கோட்பாட்டின் உறுதிப்படுத்தல்
7.1.3 ஹெர்ஷி - சேஸ் சோதனை
பின்னர், ஆல்ஃபிரட் ஹெர்ஷி மற்றும் மார்த்தா சேஸ் (1952) ஆகியோர் T2 பாக்டீரியோஃபேஜ்களுடன் நடத்திய மற்றொரு சோதனை DNA மரபணுப் பொருளாக இருப்பதற்கு ஆதாரங்களை வழங்கியது. Escherichia coli பாக்டீரியாவைப் பாதிக்கும் T2 பாக்டீரியோஃபேஜ் எனப்படும் வைரஸ் புரத உறையால் சூழப்பட்ட DNAயைக் கொண்டுள்ளது. இது ஒரு பாக்டீரியா செல்லைப் பாதிக்கும் போது, அது வெளிப்புற மேற்பரப்பில் ஒட்டிக்கொண்டு அதன் DNAயை செல்லுக்குள் செலுத்துகிறது. T2 பாக்டீரியோஃபேஜ் மற்றும் E. coli உடன் அவர்களின் தொடர் சோதனைகளில், பேஜ் துகள்களின் பெருக்கத்திற்கு எந்தக் கூறு பொறுப்பு என்பதை நிறுவுவதே நோக்கமாக இருந்தது, DNA அல்லது புரதம். எளிதாக அடையாளம் காண, $\mathrm{T} 2$ பாக்டீரியோஃபேஜ்கள் ஆரம்பத்தில் $E$. coli காலனிகளுடன் கதிரியக்க பாஸ்பரஸ் $\left({ }^{32} \mathrm{P}\right)$ மற்றும் கதிரியக்க கந்தகம் $\left({ }^{35} \mathrm{S}\right)$ ஆகியவை தனித்தனியாக உள்ள ஊடகத்தில் வளர்க்கப்பட்டன (படம் 7.3). இது ஒரு தொகுப்பு பாக்டீரியோஃபேஜ்கள் கதிரியக்க பாஸ்பரஸ் $\left({ }^{32} \mathrm{P}\right)$ உடன் மற்றொரு தொகுப்பு கதிரியக்க கந்தகம் $\left({ }^{35} \mathrm{S}\right)$ உடன் குறிக்கப்பட வழிவகுத்தது.
${ }^{35} \mathrm{S}$ மற்றும் ${ }^{32} \mathrm{P}$ குறிக்கப்பட்ட $\mathrm{T} 2$ பேஜ்கள் இப்போது குறிக்கப்படாத $E$. coli பாக்டீரியா காலனியின் இரண்டு தனி கலாச்சாரங்களுக்குள் செலுத்தப்பட்டன. பாதிப்புக்குப் பிறகு, பாக்டீரியா காலனிகள் பாக்டீரியா செல்களின் வெளிப்புறத்திலிருந்து எஞ்சியிருக்கும் எந்தப் பேஜ் மற்றும் பேஜ் பாகங்களையும் அகற்றுவதற்காக ஒரு கலக்கியில் குழப்பமடைந்தன. பின்னர் கலக்கியின் கலவையை மையவிலக்கு செய்து பாக்டீரியாக்களை (துகளில் உள்ளது) பேஜ் குப்பைகளிலிருந்து (மேல் நீர்மத்தில் உள்ளது) பிரிக்கப்பட்டது. கதிரியக்க DNA கொண்ட பேஜ்களால் பாதிக்கப்பட்ட பாக்டீரியா கலாச்சாரத்தின் துகள்கள் கதிரியக்கத்தைக் காட்டின, அதேசமயம், ${ }^{35} \mathrm{S}$ பாக்டீரியோஃபேஜால் பாதிக்கப்பட்ட மேல் நீர்மத்தில் கதிரியக்கத்தைக் காண முடிந்தது. இது புரதங்கள் பேஜிலிருந்து பாக்டீரியாக்களுக்குள் நுழையவில்லை என்பதைக் குறிக்கிறது. எனவே, பாக்டீரியா செல்லுக்குள் நுழையும் பொருள், அதாவது DNA மரபணுப் பொருளாக இருக்க முடியும் என்று முடிவு செய்யப்பட்டது.
மேற்கண்ட சோதனைகள் DNA மரபணுப் பொருளாக இருப்பதற்கு வலுவான ஆதாரங்களை வழங்கினாலும், DNA மூலக்கூறு மரபணு தகவலின் களஞ்சியம் என்பது தெளிவாக இல்லை. எர்வின் சார்காஃப், மொரிஸ் வில்கின்ஸ், ரோசாலிண்ட் ஃபிராங்க்ளின், ஜேம்ஸ் வாட்சன் மற்றும் பிரான்சிஸ் கிரிக் ஆகியோரால் செய்யப்பட்ட அடுத்தடுத்த ஆய்வுகள் DNA கட்டமைப்பின் கண்டுபிடிப்புக்கு வழிவகுத்தன, DNA எவ்வாறு பெரிய அளவிலான தகவல்களை குறியாக்கம் செய்ய முடியும் என்பதை தெளிவுபடுத்தியது (அலகு II, அத்தியாயம் 3 இல் விவரிக்கப்பட்டுள்ளது).
$\hspace{3.5cm}$பாக்டீரியோஃபேஜ்கள்

படம் 7.3: ஹெர்ஷி-சேஸ் சோதனை
7.2 புரோகேரியோடிக் மற்றும் யூகேரியோடிக் மரபணு அமைப்பு
பண்புகள் பெற்றோரிடமிருந்து சந்ததிகளுக்கு ‘மரபணு அலகு’ என பரம்பரையாக கடத்தப்படுகின்றன மற்றும் சில வைரஸ்களைத் தவிர (அங்கு மரபணுப் பொருள் RNA) அனைத்து உயிரினங்களிலும் DNA மரபணுப் பொருள் என்பது நன்கு புரிந்துகொள்ளப்பட்டுள்ளது. இது மரபணுவின் அமைப்பு பற்றிய கேள்விக்கு வழிவகுத்தது, இந்த அமைப்பு புரோகேரியோட்கள் மற்றும் யூகேரியோட்கள் இரண்டிலும் ஒரே மாதிரியாக உள்ளதா மற்றும் அது மூலக்கூறு மட்டத்தில் எவ்வாறு செயல்படுகிறது? மரபணு என்பது ஒரு குறிப்பிட்ட பண்பு அல்லது குணத்தைக் கட்டுப்படுத்தும் மரபுரிமையின் அலகு மற்றும் அலீல்கள் என அழைக்கப்படும் மாற்று வடிவங்களிலும் வெளிப்படுத்தப்படலாம். வேறுவிதமாகக் கூறினால், மரபணு என்பது RNA தொகுப்பு வழியாக பாலிபெப்டைட் சங்கிலியின் தொகுப்பு மூலம் தன்னை வெளிப்படுத்தும் DNAயின் ஒரு பகுதியாகும், இது மூலக்கூறு உயிரியலின் ‘மையக் கோட்பாடு’ என அறியப்படுகிறது.
தொடக்கத்தில், 1900 ஆம் ஆண்டில் மெண்டலின் மரபுரிமைக் கோட்பாட்டின் மறுகண்டுபிடிப்பின் அடிப்படையில், பண்புகள் அல்லது குணங்கள் மரபணுவால் கட்டுப்படுத்தப்படுகின்றன என்பது நிறுவப்பட்டது, மேலும் தாவரங்கள் மற்றும் விலங்குகள் இரண்டிலும் அடுத்தடுத்த ஆராய்ச்சிகள் ஒரு உண்மையை நிறுவின, அதாவது பண்புகள் அல்லது அம்சங்கள் சில உள்ளார்ந்த கோட்பாடுகளால் கட்டுப்படுத்தப்பட்டு ஒரு தலைமுறையிலிருந்து மற்றொன்றுக்கு அனுப்பப்படுகின்றன. பண்புகள் அல்லது குணங்களைக் கட்டுப்படுத்தும் காரணி அல்லது உள்ளார்ந்த அலகுக்கு பின்னர் 1909 ஆம் ஆண்டில் வில்ஹெல்ம் ஜோஹான்சன் ‘மரபணு’ என்ற பெயரைக் கொடுத்தார்.
மரபணுவின் இயல்பு மற்றும் செயல்பாட்டைப் புரிந்துகொள்வது இருபதாம் நூற்றாண்டின் தொடக்கத்தில் அறிவியல் சமூகத்தின் முக்கிய கவனங்களில் ஒன்றாக இருந்தது. 1930 களில் ஜார்ஜ் பீடில் மற்றும் எட்வர்ட் டேட்டம் ஆகியோரின் பணி Neurospora crassa என்ற பூஞ்சையில் ஒரு நொதியின் தொகுப்பைக் கட்டுப்படுத்த ஒரு மரபணுவின் உறவை நிறுவ உதவியது.
Neurospora crassa பூஞ்சை எளிய சர்க்கரை, கனிம உப்புகள் மற்றும் வைட்டமின் பயோட்டின் ஆகியவற்றைக் கொண்ட ஊடகத்தில் மிக எளிதாக வளர்க்கப்படும் என்ற பண்பைக் கருத்தில் கொண்டு, பீடில் மற்றும் டேட்டம் இந்த உயிரினம் மற்ற அத்தியாவசிய அமினோ அமிலங்கள் மற்றும் நைட்ரஜன் தளங்களைத் தானாகவே தொகுக்க முடியும் என்ற அடிப்படையில் சோதனை செய்தனர் (படம் 7.4). இந்தத் தொகுப்பு நொதிகளால் இடைநிலையாக நடைபெறுகிறது என்பது வெளிப்படையாகக் கருதப்பட்டது, அவை மரபணு கட்டுப்பாட்டின் கீழ் தொகுக்கப்படுகின்றன. சோதனையின் வடிவமைப்பு மிகவும் எளிமையானது, இதில் கோனிடியா, அதாவது பாலினப்பெருக்கம் செய்யாத வித்துக்கள் மரபணு மாற்றத்தைத் தூண்ட எக்ஸ்-கதிர்களால் கதிர்வீச்சு செய்யப்பட்டன (படம் 7.4). கதிர்வீச்சு செய்யப்பட்ட வித்துக்களால் உற்பத்தி செய்யப்பட்ட சந்ததிகள் சில குறிப்பிட்ட குறைந்தபட்ச ஊடகத்தில் வளர்ப்பதன் மூலம் அடையாளம் காணப்பட்டன. மரபணு மாற்றத்துடன் கூடிய இனங்களை அடையாளம் காணுவதற்காக, கதிர்வீச்சு செய்யப்பட்ட வித்துக்களின் சந்ததிகள் காட்டு வகையுடன் கலப்பினம் செய்யப்பட்டு, அடுத்தடுத்த சந்ததிகள் ஒரு குறிப்பிட்ட அமினோ அமிலம் அல்லது வைட்டமினுக்காக குறைந்தபட்ச ஊடகத்தில் (ஒரு குறிப்பிட்ட வைட்டமின் அல்லது அமினோ அமிலத்தைத் தவிர அனைத்து அமினோ அமிலங்கள், நைட்ரஜன் தளங்கள் மற்றும் வைட்டமின்களைக் கொண்ட கலாச்சார ஊடகம்) வளர்ப்பதன் மூலம் மரபணு மாற்ற இனங்கள் அடையாளம் காணப்பட்டன. அவர்களால் பல such மரபணு மாற்றங்கள் அடையாளம் காணப்பட்டன மற்றும் ஒவ்வொரு மரபணு மாற்றமும், உண்மையில், ஒரு குறிப்பிட்ட நொதியின் செயல்பாட்டின்மைக்கு விளைவாகும் என்பது மரபணு ரீதியாக நிறுவப்பட்டது.
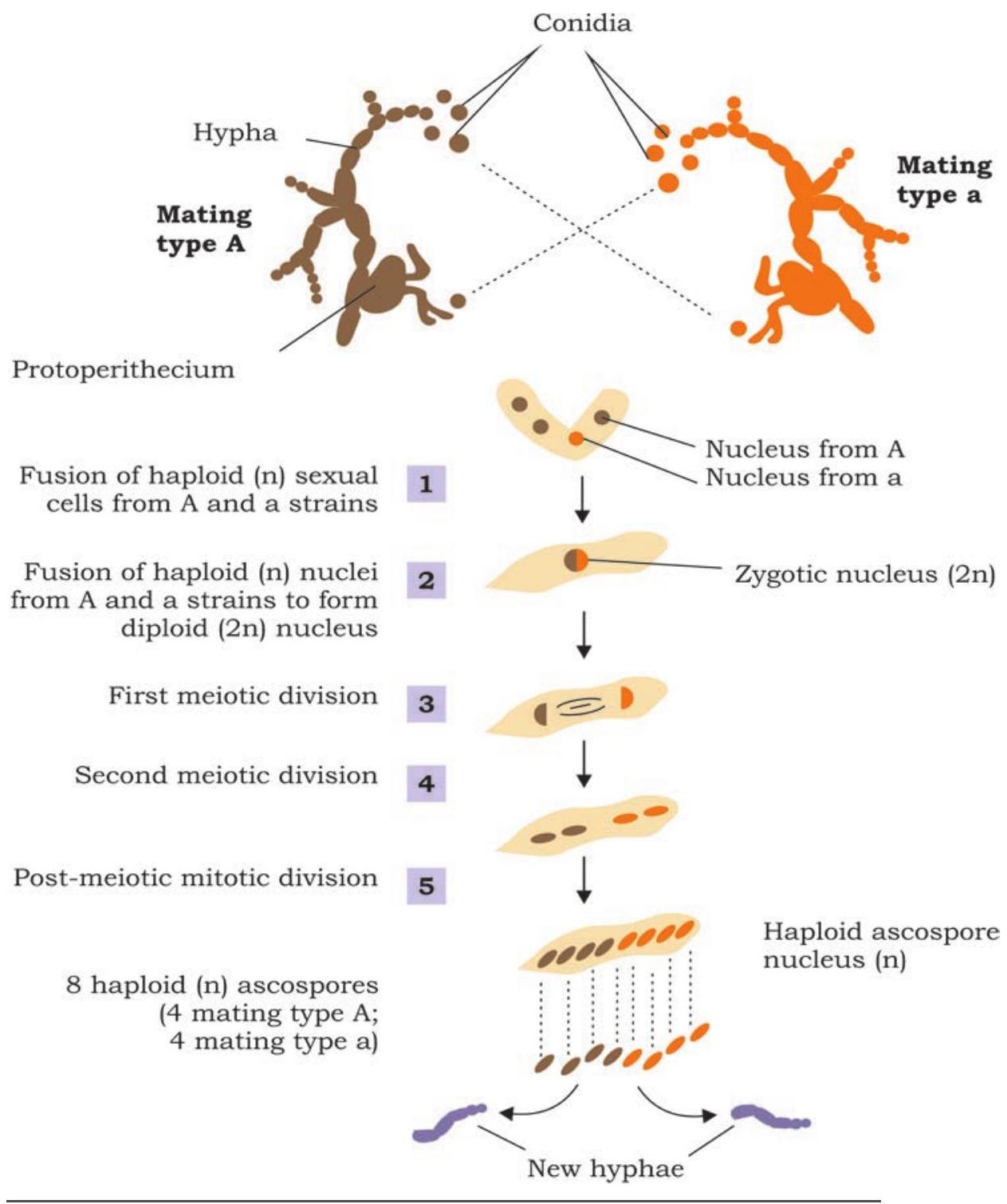
படம் 7.4: நியூரோஸ்போராவில் மரபணு மாற்றத்தைக் கண்டறிவதைக் காட்டும் சோதனை
பின்னர், அனைத்து புரதங்களும் ஒற்றை பாலிபெப்டைடால் ஆனவை அல்ல, ஆனால் ஒன்றுக்கு மேற்பட்ட பாலிபெப்டைட் சங்கிலிகளால் ஆனவை என்பது கவனிக்கப்பட்டது. ஒரு மரபணு ஒரு பாலிபெப்டைடை குறியாக்கம் செய்கிறது என்பது உண்மை; மையக் கோட்பாடும் ஒரு மரபணு ஒரு புரதம், ஒரு மரபணு ஒரு பாலிபெப்டைட் என மாற்றியமைக்கப்பட்டது.
கிட்டத்தட்ட ஒரே நேரத்தில் 1940 களின் தொடக்கத்தில், குரோமாட்டின் நாரின் உயிரணு ஆய்வுகள் மின்னணு நுண்ணோக்கி மூலம் ஓரளவு மணியின் சரம் போன்ற அமைப்பை வெளிப்படுத்தியது (படம் 7.5) மற்றும் ஒவ்வொரு மணியும் ஒரு மரபணுவைக் குறிக்கலாம் என்று உடனடியாக முடிவு செய்யப்பட்டது.

படம் 7.5: குரோமாட்டினின் சரத்தில் மணிகள்
பின்னர் ஆய்வுகள் ஒவ்வொரு மணியும் ஒரு நியூக்ளியோசோம் (ஹிஸ்டோன் எண்மத்தின் மையம் மற்றும் $146 \mathrm{~bp}$ இன் இரட்டை இழை DNA) மற்றும் இரண்டு மணிகளுக்கு இடையே உள்ள சரம், இணைப்பான் DNA ஆகியவற்றைக் கொண்டுள்ளது என்பதை வெளிப்படுத்தியது. ஒவ்வொரு நியூக்ளியோசோமும் அதன் இணைப்பான் பகுதியுடன் தோராயமாக $200 \mathrm{~bp}$ ஐ உள்ளடக்கியது என்பதும் நிறுவப்பட்டது. இது ஒரு மரபணுவாகக் கருதப்பட முடியாது, ஏனெனில் பல சந்தர்ப்பங்களில் மரபணுக்களின் அளவு 200 நியூக்ளியோடைடுகளை விட மிகப் பெரியதாக இருக்க வேண்டும். எளிய காரணம் என்னவென்றால், பல புரதங்களுக்கு 100 க்கும் மேற்பட்ட அமினோ அமில எச்சங்கள் உள்ளன மற்றும் அவற்றின் தொடர்புடைய கட்டுப்பாட்டு மரபணுக்கள் மூன்று மடங்குக்குக் குறையாது (கோடானின் மும்மைத் தன்மையின் அடிப்படையில்).
ஒரு மரபணு என்பது குறிப்பிட்ட ஊக்குவிப்பாளர் பகுதியுடன் கூடிய DNAயின் பகுதியாகும், அங்கு RNA பாலிமரேஸ் பிணைக்க முடியும் மற்றும் mRNAயை படியெடுக்க முடியும் என்பது இப்போது தெளிவாகியுள்ளது. படியெடுக்கப்பட்ட mRNA பின்னர் மொழிபெயர்ப்பு செயல்முறையில் ஈடுபடுகிறது. வைரஸ் முதல் பாக்டீரியா, தாவரங்கள் மற்றும் விலங்குகள் வரை அனைத்து உயிரினங்களுக்கும் இந்த பொறிமுறை ஒன்றே. ஒரு வைரஸ், பாக்டீரியா அல்லது உயர் உயிரினத்தின் மரபணுக்கள் அவற்றின் கட்டமைப்பு மற்றும் செயல்பாட்டில் ஒரே மாதிரியானவையா? ஒரு முழுமையான குரோமோசோம்களின் தொகுப்பில் உள்ள மொத்த DNA உள்ளடக்கம் (அனைத்து மரபணுக்கள் மற்றும் DNAயின் பிற பகுதிகளையும் உள்ளடக்கியது) மரபணுத்தொகை என்று அழைக்கப்படுகிறது. வைரஸ்கள் அல்லது பாக்டீரியாக்களின் விஷயத்தில் நாம் பார்ப்பது போல, மரபணுத்தொகையின் அளவு யூகேரியோடிக் மரபணுத்தொகையுடன் ஒப்பிடும்போது ஒப்பீட்டளவில் மிகச் சிறியது. யூகேரியோடிக் மரபணுத்தொகைகள் புரோகேரியோடிக் மரபணுத்தொகைகளை விட மிகவும் சிக்கலானவை. தாவர மரபணுத்தொகைகள் வேறு எந்த யூகேரியோடிக் மரபணுத்தொகைகளை விட இன்னும் சிக்கலானவை. வாழும் உயிரினங்களின் பல்வேறு குழுக்களில் மரபணுத்தொகையின் மதிப்பிடப்பட்ட அளவு படம் 7.6 இல் காட்டப்பட்டுள்ளது.
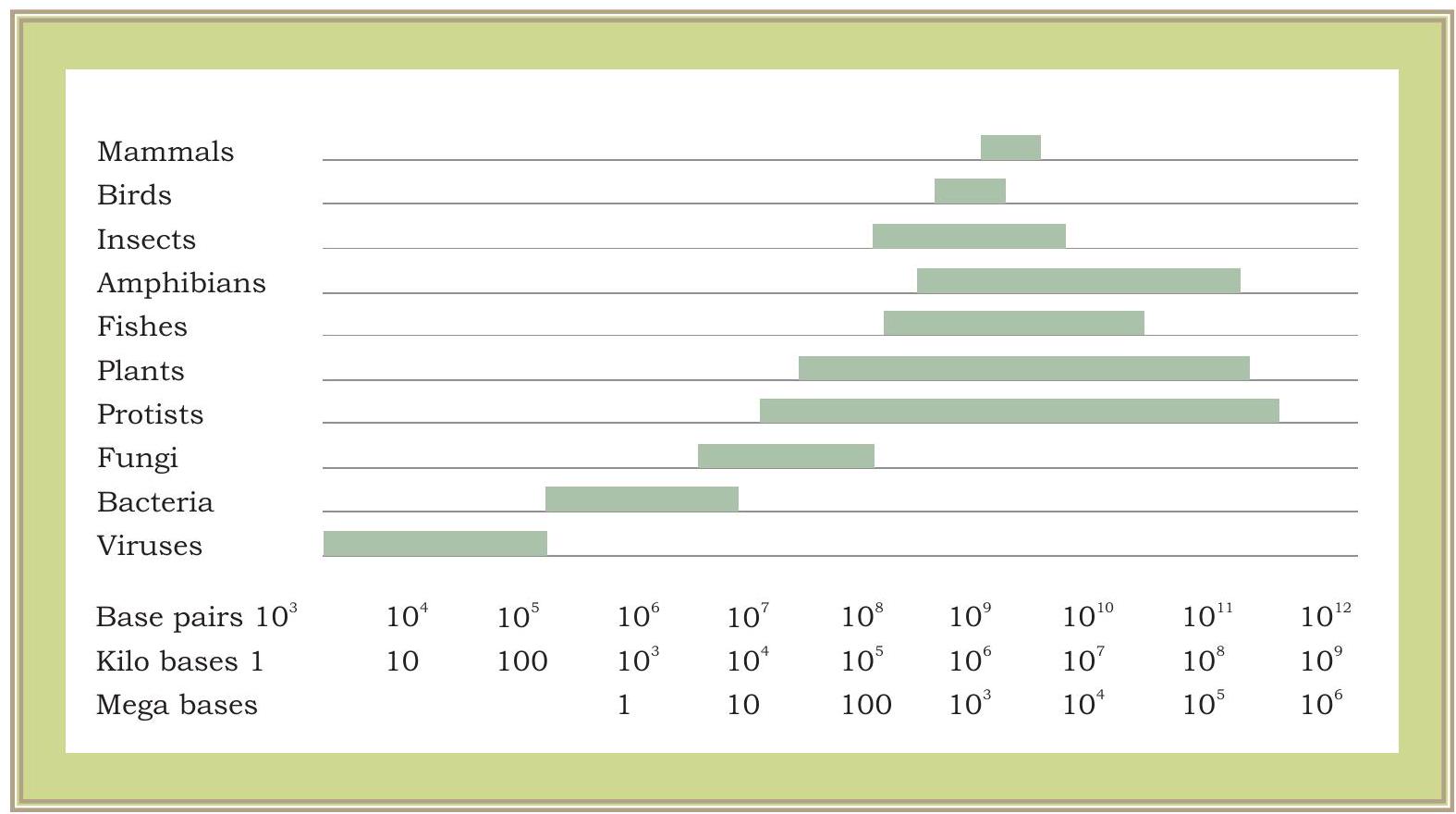
படம் 7.6: வாழும் உயிரினங்களின் பல்வேறு குழுக்களில் மரபணுத்தொகை அளவு மாறுபாடுகள். மரபணுத்தொகை அளவுகள் ஆயிரக்கணக்கான நியூக்ளியோடைட் இணைகளில் அளவிடப்படுகின்றன, அதாவது $1000 \mathrm{bp}=1$ கிலோபேஸ் (kb) மற்றும் 1000,000 $\mathrm{bp}=1000 \mathrm{~Kb}=1$ மெகாபேஸ் (Mb)
பெரும்பாலான யூகேரியோட்களில், மரபணுத்தொகையின் ஒரு பெரிய பகுதி வெளிப்படுத்தப்படாமல், குறியீடு அல்லாத வரிசையாகவே இருக்கும். யூகேரியோடிக் மரபணு வெளிப்பாட்டில், பாலிபெப்டைட் சங்கிலி தொகுப்பு செயல்பாட்டில் ஈடுபடும் செயலில் உள்ள mRNAயின் அளவு முதன்மை படியெடுப்பை விட மிகச் சிறியது என்பதும் கவனிக்கப்பட்டுள்ளது. உண்மையில், பல யூகேரியோடிக் மரபணுக்கள், எ.கா., ஹீமோகுளோபினின் $\beta$-குளோபின் மரபணு படியெடுத்த பிறகு இணைப்பு செயல்முறையை அனுபவிக்கிறது, இதில் முதன்மை படியெடுப்பின் சில இடைப்பட்ட பகுதிகள் வெட்டி எடுக்கப்படுகின்றன (இன்ட்ரான்கள்) மற்றும் மீதமுள்ள பகுதிகள் (எக்ஸான்கள்) RNA படியெடுப்பு mRNA உருவாக்க இணைக்கப்படுகின்றன.
ஒரு யூகேரியோடிக் செல்லுக்கு இரண்டு வகையான மரபணுத்தொகைகள் உள்ளன: (i) அணு மரபணுத்தொகை மற்றும் (ii) உறுப்பு மரபணுத்தொகை.
அணு மரபணுத்தொகை
பெரும்பாலான DNA கருவில் காணப்படுகிறது மற்றும் அணு DNA என அறியப்படுகிறது. புரோகேரியோட்களில், பெரும்பாலான மரபணுத்தொகை குறியீட்டு DNA வரிசைகளால் ஆனது, அதேசமயம் யூகேரியோடிக் மரபணுத்தொகைகளில் குறியீட்டு பகுதிகள் மொத்த மரபணுத்தொகையின் ஒப்பீட்டளவில் மிகச் சிறிய பகுதியை உருவாக்குகின்றன. எடுத்துக்காட்டாக, மனித மரபணுத்தொகையின் அளவு தோராயமாக $3,000 \mathrm{Mb}$ அல்லது 3 பில்லியன் ஜோடி தளங்க